Phylogeny of the beetles |
Text © Christoph Benisch, 2010
|
|
|
1. Introduction |
|
In evolutionary terms, beetles are an old order (see Fig. 1a). The oldest fossil beetles are known from the Permian, a long time before the dinosaur ruled the world.
Since these days, the beetles evolved into the most speciose order in the animal kingdom. Worldwide, more than 350,000 species have been scientifically
described. That is, 40% of all scientifically described insects and 25% of all described animals are beetles. Even today, every year a large number
of new species is discovered. There are estimates, that up to 8 mio. beetle species exist worldwide.
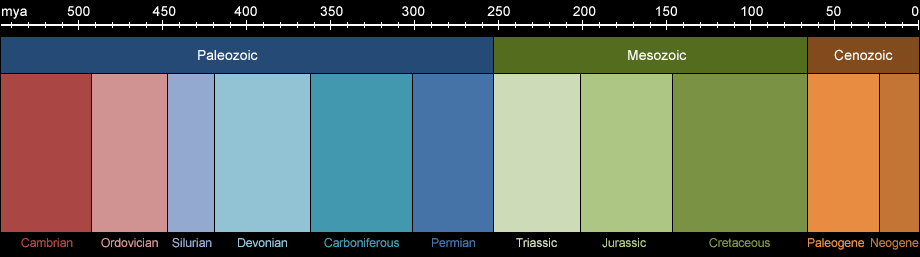
Fig. 1a: Geological eras from the Paleozoic to today. Click to display selected animal groups.
Global climate conditions changed several times during the history of the earth. They can be reconstructed until about 2.3 billion years ago by sediment analysis.
During the Phanerozoic several ice ages occurred, characterized by a complete glaciation of the polar caps. From a geological point of view, today we live in an ice age as well.
However, this is not at all the "normal" climate condition of the earth. On the contrary, during the last 550 mio. years acryogenic warm periods prevailed for around 80% of the time,
with the polar caps completely free of ice. Fig. 1b shows the global average temperature in a strongly simplified form.
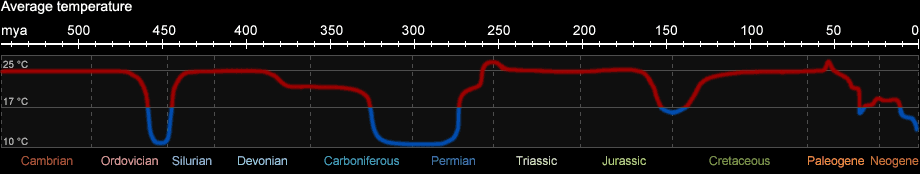
Fig. 1b: Climate conditions during the Phanerozoic (after C. R. Scotese, Paleomap, 2008)
The silurian-ordovician ice age began about 450 mio. years ago and was confined to what is today North Africa and South America. Due to strong cooling of the oceans
it was accompanied by a mass extinction of species (see Fig. 1c). The short ice age was followed by a warm period lasting from the upper Silurian until the middle Carboniferous.
In the later part of the Carboniferous, the so called Permo-Carboniferous ice age began. The glaciation was mainly confined to the southern hemisphere and the period is often
referred to as "Gondwana glaciation". The last ice age (also called quaternary ice age) began around 2.6 mio. years ago and lasts until today. It is characterized by
rapid changes between cold periods (glacials) and intermittent warm periods (interglacials).
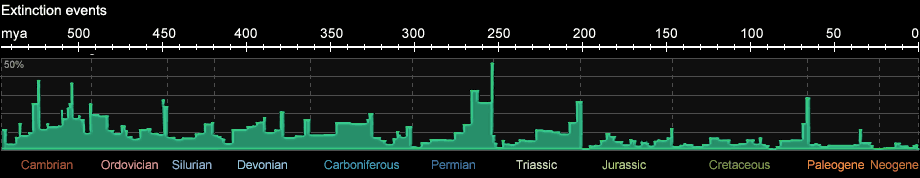
Fig. 1c: Extinction events during the Phanerozoic (after R. A. Rohde and R. A. Muller, 2005)
|
2.0 Permian (299-250 mya) |
|
During the Permian the land masses aggregated to a single supercontinent, called Pangea (see Fig. 2). It was formed by the collision of Gondwana (today's South America,
Africa, Antarctica, Australia, Madagascar and India) with Laurussia (today's Europe and North America) and Siberia (today's middle-siberian mountains). During the collision
the Ural was folded. The supercontinent was surrounded by a single ocean, the so called Panthalassic Ocean. The ocean in the eastern bay of Pangea is called Tethys Ocean.
Major parts of today's Europe were covered by a salty inland sea (Zechstein sea). Its basin dried up and was flooded several times during the Permian.
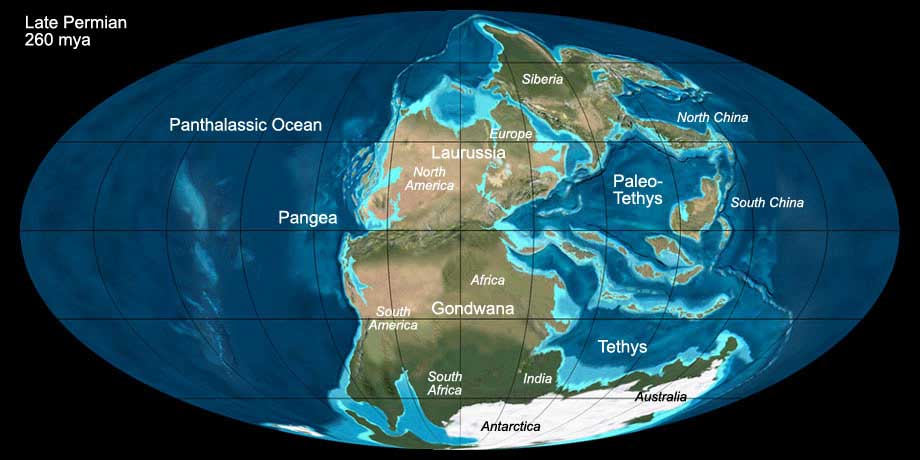
Fig. 2: The World during the Upper Permian (260 mya). Click to show/hide labels and climate. © Ron Blakey, use with written permission.
At the beginning of the Permian climate was characterized by the Permo-Carboniferous ice age. The polar caps were covered by massive glaciers, major parts of Gondwana
were strongly glaciated as well. In the tropical regions huge forest swamps grew up. During the Middle Permian the climate became warmer and drier and the glaciers retreated.
The inland was characterized by continental climate with strong seasonal fluctuations in temperature and rainy and dry seasons.
| Period |
Epoch |
Stage |
Age (mya) |
| Permian |
Lopingian |
Changhsingian |
251,0 - 253,8 |
| Wuchiapingian |
253,8 - 260,4 |
| Guadalupian |
Capitanian |
260,4 - 265,8 |
| Wordian |
265,8 - 268,0 |
| Roadian |
268,0 - 270,6 |
| Cisuralian |
Kungurian |
270,6 - 275,6 |
| Artinskian |
275,6 - 284,4 |
| Sakmarian |
284,4 - 294,6 |
| Asselian |
294,6 - 299,0 |
The oldest beetle fossils are from the Lower Permian (Asselian stage) and are approximately 300 mio. years old, for instance in the red slate fossil beds of
Niedermoschel near Mainz, Germany1. Further fossils have been found in Obora, Czechia2 (Artinskian stage)
and Tshekarda in the Ural mountains, Russia3,4 (Kungurian stage). Interestingly, contrary to the situation in Europe and
Asia, there are so far only very few fossil beetles known from the Artinskian stage of North America, although both continents had been united to Laurussia.
The first discoveries from North America were made in the Wellington formation of Oklahoma and were published in 2005 and 20085,6.
In the upper stages of the Middle Permian in Europe and Western Asia, beetles have been recorded rather infrequently. In 2003 PONOMARENKO
published the first fossil beetle record from the Tartarian stage (the regional equivalent of the Capitanian stage)7.
In the Upper Permian beetles appeared on all continents, especially Laurussia and Siberia, later in Gondwana as well.
For instance, discoveries of beetle fossils were made in South Africa8, South America, Australia and Eastern Europe.
In most cases, the fossils consist of (mostly isolated) elytra, which had developed during the Permian by transmutation of the membranous forewings into sheathed elytra.
This development was finalized by the end of the Permian.
Fig. 3: Elytron of Permocoleus wellingtonensis, Wellington formation, Oklahoma, USA (Lower Permian, Artinskian stage). Click to enlarge.
© Roy Beckemeyer, use with written permission.
At the end of the Permian, the biggest mass extinction in Earth's history took place (the so called P-Tr extinction event): 95% of all marine species
and 66% of all terrestrial species became extinct. Also 30% of all insect species became extinct, the only mass extinction of insects in Earth's history until today.
The reasons for the mass extinction are not fully clarified. Possible mechanisms include volcanism, a meteorite impact, a sudden release of methane hydrates from the sea floor or
a cosmic gamma-ray burst.
|
2.1 Triassic (251-199 mya) |
|
Pangea had already started to break apart in the Upper Permian and rift valleys formed between North America and Africa. During the Triassic the Tethys Sea protruded
into Pangea towards the west, and in the Upper Triassic rift valleys formed between North America and Europe, followed by marine ingression.
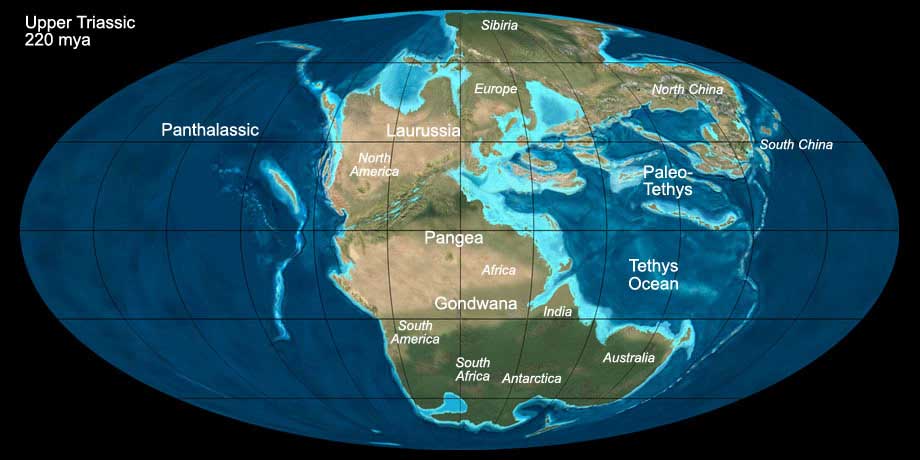
Fig. 4: The world during the Upper Triassic (220 mya). Click to show/hide labels and climate. © Ron Blakey, use with written permission.
Throughout the whole Triassic, climate was warm to hot and mostly dry, in the inland of Pangea even arid and with a strong continental character with
very hot summers and cold winters. The sea level was rather low during the Triassic, contributing to the warm climate. The polar caps were free of ice.
| Period |
Epoch |
Stage |
Age (mya) |
| Triassic |
Upper Triassic |
Rhaetian |
199,6 - 203,6 |
| Norian |
203,6 - 216,5 |
| Karnian |
216,5 - 228,7 |
| Middle Triassic |
Ladinian |
228,7 - 237,0 |
| Anisian |
237,0 - 245,9 |
| Lower Triassic |
Olenekian |
245,9 - 249,5 |
| Induan |
249,5 - 251,0 |
As a consequence of the mass extinction at the border of Permian and Triassic, there is only little fossil record of insects including beetles from the Lower Triassic
9. However, there are a few exemptions, e.g. in Eastern Europe: At the Babiy Kamen site10
in the Kuznetsk Basin (Induan- and Olenekian stage) numerous beetle fossils were discovered, even entire specimen of the infraorders
Archostemata (Ademosynidae, Schizocoleidae), Adephaga (Triaplidae, Trachypachidae) and Polyphaga
(Hydrophilidae, Byrrhidae, Elateroidea) in perfectly preserved condition.
However, species from the families Cupedidae and Schizophoroidae are not present at this site, whereas they dominate at other fossil sites from the Lower Triassic.
Further records are known from Khey-Yaga11 in the Korotaikha Basin (Olenekian and Anisian stage).
Since the Middle Triassic, mycetophagous Cupedidae appear in the fossil record. In the stages of the Upper Triassic representatives of the algivorous Triaplidae
and Hydrophilidae are present, as well as predatory water beetles. The first primitive weevils appear (Obrienidae), as well as the first representatives
of the rove beetles (Staphylinidae), which show no marked difference in physique compared to recent species.
 Fig. 5: Elytron of Argentinosyne duraznoensis (Schizocoleidae), Potrerillos Formation, Mendoza Province, Argentina (Early Upper Triassic, Carnian stage).
© Oscar F. Gallego, use with written permission.
Fig. 5: Elytron of Argentinosyne duraznoensis (Schizocoleidae), Potrerillos Formation, Mendoza Province, Argentina (Early Upper Triassic, Carnian stage).
© Oscar F. Gallego, use with written permission.
|
2.2 Jurassic (199-145 mya) |
|
During the Jurassic the supercontinent Pangea broke apart into a northern part (Laurasia) and a southern part (Gondwana), a process that had started in the Triassic already.
The northern Atlantic Ocean was still very narrow and the southern Atlantic Ocean opened only towards the end of the Jurassic.
In North America the Rocky Mountains started to fold. Antarctica was separated from Africa and South America by a rift valley and India started to drift northwards.
The sea level was somewhat higher than today, and major parts of Europe and North America were covered by a shallow tropical sea with a great biodiversity.
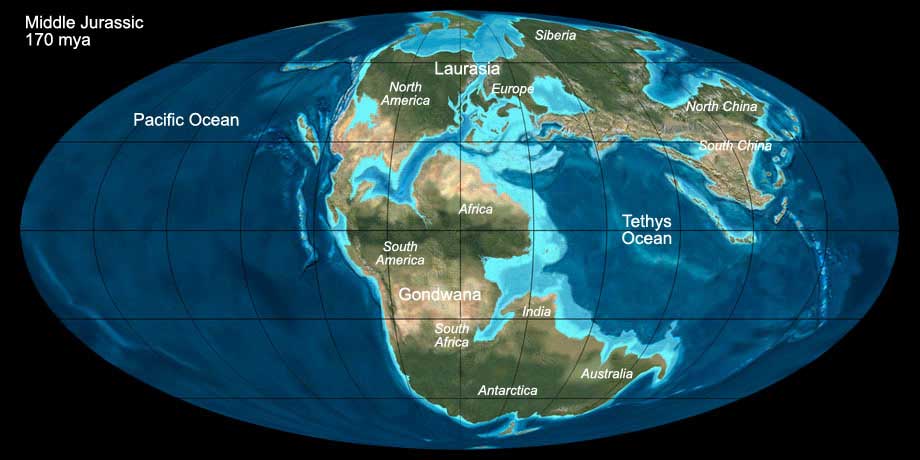
Fig. 6: The world during the Middle Jurassic (170 mya). Click to show/hide labels and climate. © Ron Blakey, use with written permission.
The global climate during the Jurassic was warm and humid. Similar to the Triassic, there were no larger landmasses situated near the polar caps and consequently, no
inland ice sheets existed during the Jurassic. Although some areas of North and South America and Africa stayed arid, large parts of the continental landmasses were lush.
The northern (laurasian) and the southern (gondwanian) fauna differed considerably in the Early Jurassic. Later it became more intercontinental and many species started to spread globally.
| Period |
Epoch |
Stage |
Age (mya) |
| Jurassic |
Upper Jurassic |
Tithonian |
145,4 - 150,8 |
| Kimmeridgian |
150,8 - 155,7 |
| Oxfordian |
155,7 - 161,2 |
| Middle Jurassic |
Callovian |
161,2 - 164,7 |
| Bathonian |
164,7 - 167,7 |
| Bajocian |
167,7 - 171,6 |
| Aalenian |
171,6 - 175,6 |
| Lower Jurassic |
Toarcian |
175,6 - 183,0 |
| Pliensbachian |
183,0 - 189,6 |
| Sinemurian |
189,6 - 196,5 |
| Hettangian |
196,5 - 199,6 |
There are more than 150 important sites with beetle fossils from the Jurassic, the majority being situated in (Eastern) Europe and North Asia.
In North America and especially in South America and Africa the number of Jurassic sites is smaller and the sites have not been exhaustively investigated yet.
Outstanding fossil sites include Solnhofen12 (Tithonian stage) in Upper Bavaria, Germany, Karatau13 in South Kazakhstan (Oxfordian or
Kimmeridgian stage), the Yixian formation in Liaoning14, North China (Tithonian-Berriasian stage,
contains the Jehol biota) as well as the Jiulongshan formation15,16 (Aalenian or Bajocian stage, contains the Yanliao biota)
and further fossil sites17 in Mongolia. In North America there are only a few sites with fossil records of insects from the Jurassic, namely the
shell limestone deposits in the Hartford basin, the Deerfield basin and the Newark basin18 (Hettangian-Sinemurian stage).
The beetles from the Lower Jurassic are similar to the species known from the Upper Triassic. Xylophagous Cupedidae only appear in warm zones.
Towards the Upper Jurassic the fraction of the Cupedidae decreased and at the same time the diversity of the early phytophagous species increased. Most of the recent phytophagous beetle
species feed on flowering plants or angiosperms (Angiospermae). It is believed that the increase in diversity of the angiosperms also influenced the diversity of the phytophagous species,
which doubled during the Middle Jurassic. However, recently doubts have been raised since the increase of the number of beetle families during the Cretaceous does not correlate
with the increase of the number of angiosperm species19.
In the Upper Jurassic numerous primitive weevils20 (Curculionoidea) and click beetles21,22
(Elateroidea) appeared. Also first jewel beetles23-25 (Buprestidae) are present, however, they were rather rare until the Cretaceous.
Fig. 7: 1a,b: Paradesmatus baiae; 6a,b: Paradesmatus ponomarenkoi; 9: Protagrypnus robustus; Jiulongshan formation, Ningcheng Province, Inner Mongolia, China (Middle Jurassic, Aalenian or Bajocian stage). Click to enlarge.
© Ren Dong and Huali Chang, use with written permission.
|
2.3 Cretaceous (145-65 mya) |
|
The Cretaceous is the youngest period of the Mesozoic and lasted 80 mio. years (from 145 until 65 mya).
During the Cretaceous the former supercontinent Pangea finally broke apart. The landmasses of Africa and South America - still connected during the Lower Cretaceous - separated
and opened the South Atlantic Ocean. At the beginning of the Turonian it was connected to the North Atlantic.
Australia and India started to drift northwards apart from Antarctica. Madagascar and India stayed connected for the time being.
Major parts of North America and Europe were covered by a shallow sea, which retreated only at the end of the Creatceous.
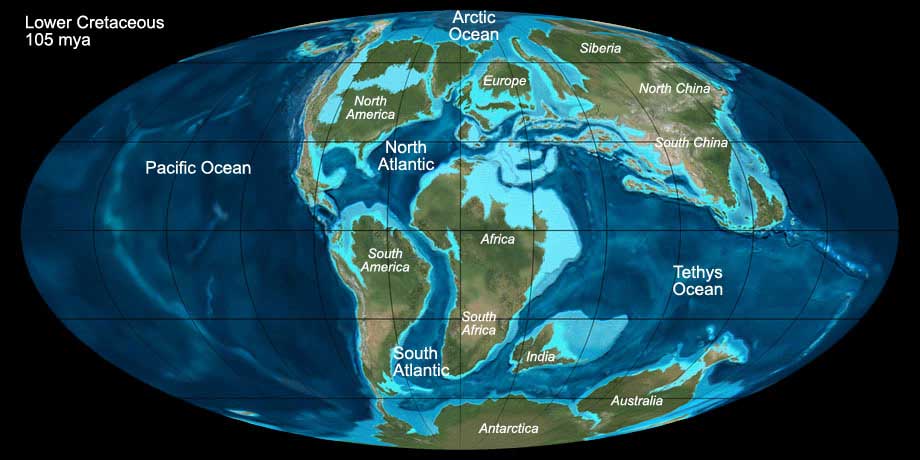
Fig. 8: The world during the late Lower Cretaceous (105 mya). Click to show/hide labels and climate. © Ron Blakey, use with written permission.
At the beginning of the Cretaceous, in the Barremian stage, the climate continued to cool, a process that gad started in the Upper Jurassic already. Presumably there was
snow in higher altitudes, however, glaciation outside higher altitudes did not take place. At the end of the Berriasian, the temperatures increased again and stayed relatively
constant until the end of the Cretaceous. The CO2 concentration in the atmosphere increased due to volcanism and contributed to the global warming.
Near the polar caps the climate was cool temperate, and dinosaurs appeared up to the 15° latitude from the south pole. The flat global temperature gradient caused only little
wind, and the oceans were less agitated. Deep ocean anoxia obviously often occurred, as indicated by clay slate sediments.
| Period |
Epoch |
Stage |
Age (mya) |
| Cretaceous |
Upper Cretaceous |
Maastrichtian |
65,5 - 70,6 |
| Campanian |
70,6 - 83,5 |
| Santonian |
83,5 - 85,8 |
| Coniacian |
85,8 - 88,6 |
| Turonian |
88,6 - 93,5 |
| Cenomanian |
93,5 - 99,6 |
| Lower Cretaceous |
Albian |
99,6 - 112,0 |
| Aptian |
112,0 - 125,0 |
| Barremian |
125,0 - 130,0 |
| Hauterivian |
130,0 - 133,9 |
| Valanginian |
133,9 - 140,2 |
| Berriasian |
140,2 - 145,5 |
There is a large number of important fossil sites worldwide containing beetles from the Cretaceous. Most of of them are located in Europe and Asia and
belong to the temperate climate zone during the Cretaceous. A few of the fossil sites mentioned in the chapter Jurassic also shed some light on the early cretaceous beetle fauna,
e.g. the Yixian formation in Liaoning14, North China (Tithonian-Berriasian stage). Further important sites from the Lower Cretaceous
include the Crato Fossil Beds26 in the Araripe basin in the Federal State Ceará, North Brazil
(Aptian-Albian stage) as well as overlying Santana formation (Cenomanian stage). The latter was situated near the Paleoequator.
In Spain there are important sites near Montsec (Hauterivian-Barremian stage) and Las Hoyas27 (Barremian stage).
In Australia the Koonwarra fossil beds (Aptian stage) of the Korumburra group, South Gippsland, Victoria is noteworthy.
Important fossil sites from the Upper Cretaceous are Kzyl-Dzhar (Turonian stage) in South Kazakhstan and Arkagala (Turonian stage) in Russia.
During the Cretaceous the diversity of the Cupedidae and Archostemata decreased considerably. Predatory ground beetles (Carabidae) and rove beetles
(Staphylinidae) showed a different distribution pattern: Whereas the Carabidae predominantly occurred in the warm regions, the Staphylinidae preferred the
zones with temperate climate. Likewise predatory Cleroidea and Cucujoidea, hunted their prey under the bark of trees.
Click beetles (Elateridae) were common in the warm regions. Together with the jewel beetles (Buprestidae)28,
whose diversity increased rapidly during the Cretaceous, they were the primary consumers of wood29.
Longhorn beetles (Cerambycidae) were rather rare and their diversity increased only towards the end of the Upper Cretaceous.
 Fig. 9: Left: Drawing of Cratomacer immersus (Nemonychidae), Santana Formation, Crato Member, Ceará, North Brazil (Aptian-Albian stage),
redrawn after V. V. Zherikhin and V. G. Gratshev, Paleontological Journal, 2004, 38(5), 530. Right: Alexcarabus megagnathus (Carabidae),
Santana Formation, Crato Member (see above), © Rafael G. Martins-Neto, use with written permission.
Fig. 9: Left: Drawing of Cratomacer immersus (Nemonychidae), Santana Formation, Crato Member, Ceará, North Brazil (Aptian-Albian stage),
redrawn after V. V. Zherikhin and V. G. Gratshev, Paleontological Journal, 2004, 38(5), 530. Right: Alexcarabus megagnathus (Carabidae),
Santana Formation, Crato Member (see above), © Rafael G. Martins-Neto, use with written permission.
The first scarab beetles (Scarabaeidae) appeared in the Upper Jurassic. They were not yet coprophagous, but presumably fed on decaying wood infested with fungi.
The first coprophagous beetles have been recorded from the Upper Cretaceous (Campanian stage)30, and lived on the excrements of herbivorous dinosaurs.
There is still a discussion, whether the coprophagy the beetles was always tied to mammal excrements during its development in the Upper Cretaceous31.
Among the water beetles the first species with an adaption of both larvae and adults to the aquatic lifestyle are found. Whirligig beetles (Gyrinidae)
were moderately diverse, early Dytiscid beetles less. The most widespread were the Coptoclavidae, which preyed on aquatic fly larvae.
Feeding traces of phytophagous leaf beetles (Chrysomelidae) have been recorded from the Upper Cretaceous for the first time32. In the 2nd half
of the Upper Cretaceous the Donaciinae dwelled on aquatic plants. Weevils (Curculionidae) appeared towards the end of the Lower Cretaceous.
The fossil records of beetles are predominantly from Asia, a few from Europe33 and Brazil34 as well.
At the end of the Cretaceous 65 mio. years ago, at to so called K-T-boundary (from German: Kreide-Tertiär, "Tertiär" being to former name for the Paleogene) a big mass extinction took place,
that particularly ended the reign of the dinosaur. It is assumed that one or several meteorite impacts and increased volcanism during a period of about 1 mio. years
caused serious environmental changes. The theory of a meteorite impact is supported by the iridium and chromium anomaly in sediments at the K-T-boundary.
However, beetles species seemed to be little affected. For example, in Eastern Russia there are deposits on both sites of the K-T-boundary with a similar diversity.
|
2.4 Paleogene (65-23 mya) |
|
The Paleogene comprises the first part of the Cenozoic and lasted from 65 to 23 mio. years ago. During this time the continents assumed their modern shapes.
The fragments of Gondwana (South America, Africa, India and Australia) began to drift northwards. The collision of India with the eurasian landmass led to the
folding of the Himalaya. Similarly, the Alps were folded in Central Europe by the collision of the African plate with Europe. South America's drift was less pronounced
and the "collision" with North America was cushioned by a system of smaller plates in the Caribbean region, preventing the folding of mountains.
A land bridge between North America and South America did not yet exist. The Atlantic Ocean continued to widen during the Paleogene. In the North, the last land bridge between
North America and Europe broke up during the Eocene.
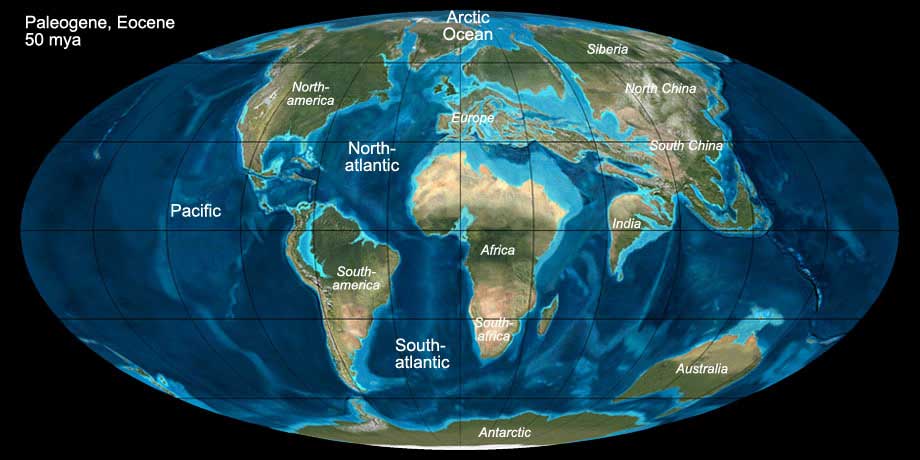
Fig. 10: The world during the Paleogene (Eocene, 50 mya). Click to show/hide labels and climate. © Ron Blakey, use with written permission.
| Period |
Epoch |
Stage |
Age (mya) |
| Paleogene |
Oligocene |
Chattian |
28,4 - 23,0 |
| Rupelian |
33,9 - 28,4 |
| Eocene |
Priabonian |
37,2 - 33,9 |
| Bartonian |
40,4 - 37,2 |
| Lutetian |
48,6 - 40,4 |
| Ypresian |
55,8 - 48,6 |
| Paleocene |
Thanetian |
58,7 - 55,8 |
| Seelandian |
61,1 - 58,7 |
| Danian |
65,5 - 61,1 |
Climate during the Paleogene was warm and tropical as most time during the Mesozoic. At the beginning, in the Paleocene, climate was drier and cooler than
in the preceding Cretaceous. However, temperature strongly increased during the Eocene and subtropical vegetation spread up to Greenland and Patagonia.
The climate near the poles was cool temperate, in Europe, North America, Australia and the southern part of South America warm temperate.
Near the equator there was tropical climate, flanked by hot and arid zones in the north and the south. In the Oligocene, global cooling started.
Antarctica was covered by an ice sheet and subsequently, sea levels dropped. Except an intermittent warm period during the late Oligocene, global
cooling continued and finally led to the Pleistocene ice age.
There are numerous fossil beetles known from the Paleogene. The beetle fauna of the Paleocene (61-56 mya) is comparatively poorly investigated.
In contrast, the knowledge on the Eocene beetle fauna is very good. The reason is the occurrence of fossil insects in amber and cay slate sediments.
Amber is fossilized tree resin, that means it consists of fossilized organic compounds, not minerals. Ambers are distinguished by location, age and species of the resin producing plant.
For the research on the Oligocene beetle fauna, Baltic35,36 and Dominican amber37 is most important.
 |
 |
 |
|
Fig. 11: Left: Sucinoptinus sucini, Baltic amber, Upper Eocene, © Francesco Vitali. Center: Curculionidae sp., Dominican amber, Late Oligocene,
© Amberica West, USA. Right: Platypodidae sp., Dominican amber, Late Oligocene, © Amberica West, USA. Use with written permission.
|
Baltic amber (succinite) is mainly found on the coasts of the North Sea and the Baltic Sea and originates from the Eocene (mainly Lutetian, 50-40 mya).
According to today's view, it was formed by a close relative of the recent Golden Larch (Pseudolarix).
Dominican amber originates from the Eocene to the Lower Miocene, the resin was formed by Hymenaea protera. Dominican amber is well known for its
transparency and a high number of fossil inclusions. Further interesting beetle fossils from the Paleogene were recorded from British Columbia,
Canada38. Generally, representatives of the beetle families Elateridae, Anobiidae, Scirtidae and Staphylinidae are predominant in amber.
Oil shale sediments are another source of beetle fossils. They formed in deep water zones by the decomposition of organic matter under anoxic conditions.
In this way, decay was prevented and the decomposition of organic material finally led to hydrocarbons.
Especially important is the Messel Pit Fossil Site near Darmstadt, Germany, an UNESCO World Heritage site since 1995. Its oil shales formed 47 mio. years ago
during a period of 1.5 mio. years in a 300 m deep maar. Besides excellently preserved birds, mammals, reptiles and fishes, also beetle fossils have been discovered
in the Messel pit. Even today, they show their original splendid colors39 (see Fig. 12). Jewel beetles (Buprestidae) and click beetles (Elateridae)
are among the more common families recorded from oil shales, whereas discoveries of fossil longhorn beetles (Cerambycidae) considerably scarcer.
|
2.5 Neogene (23 mya until today) |
|
During the Neogene the continents finally assumed their today's position. The South American continent drifted to the west towards the subduction zone in the Pacific.
During this process the Andes were folded. During the Pliocene (5 mya) the land bridge between South America and North America was formed, and the fauna exchange started.
The formation of this land bridge also had an impact on global climate. The Indian subcontinent continued it collision with Asia, but added an westward movement as well,
leading to the folding of the Caucasus. The folding of the Himalaya continues until today.
The collision of Africa with Europe and the rise of the lithosphere under the Alborán Sea (westernmost Mediterranean) lead to the separation of the Mediterranean from the Atlantic Ocean.
During this period, that lasted 600.000 years from 6 until 5.3 mya, the Mediterranean desiccated nearly completely (Messinian salinity crisis). Only at the end of this
period the desiccated basin was flooded through a narrow canal near Gibraltar, according to today's view quickly, but without catastrophic effects.
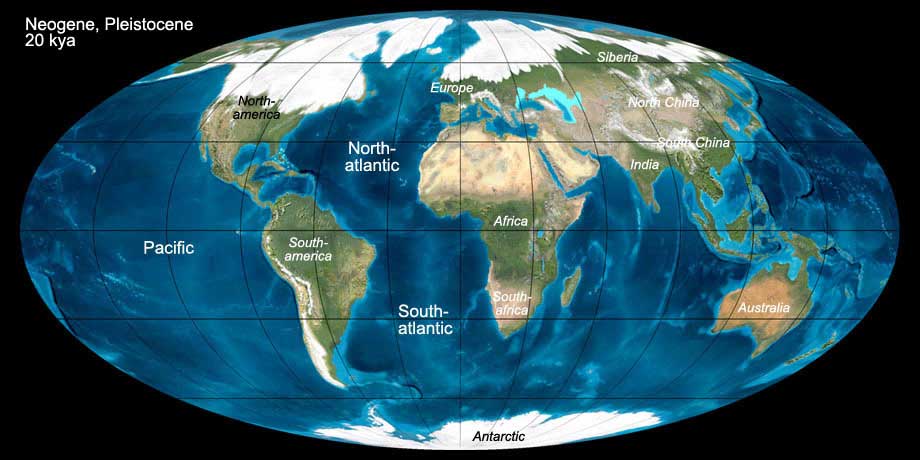
Fig. 13: The world during the Neogene (Pleistocene, 20 kya). Click to show/hide labels and climate. © Ron Blakey, use with written permission.
| Period |
Epoch |
Stage |
Age (mya) |
| Neogene |
Holocene |
Holocene |
0,0117 - 0 |
| Pleistocene |
Upper Pleistocene
(Tarantian) |
0,126 - 0,0117 |
Middle Pleistocene
(Ionian) |
0,781 - 0,126 |
Lower Pleistocene
(Calabrian) |
1,806 - 0,781 |
| Gelasian |
2,588 - 1,806 |
| Pliocene |
Piacenzian |
3,600 - 2,588 |
| Zanclean |
5,332 - 3,600 |
| Miocene |
Messinian |
7,246 - 5,332 |
| Tortonian |
11,608 - 7,246 |
| Serravallian |
13,82 - 11,608 |
| Langhian |
15,97 - 13,82 |
| Burdigalian |
20,43 - 15,97 |
| Aquitanian |
23,03 - 20,43 |
The Neogene was a period of global cooling, which finally led to the Pleistocene ice age. At the beginning of the Miocene, temperatures in the northern hemisphere
initially were still temperate. However, by the formation of the land bridge between South America and North America, the warm ocean current was cut off and the polar
caps cooled down dramatically. During the Gelasian period ice sheets began to form both in the Arctic and Antarctic region. This marked the starting point of a new ice age
which continues until today, with glacial cycles and intermittent warmer periods (interglacials). During the glacials the continental glaciers pushed to the 40th parallel
in some regions and covered major parts of North America, Europe and Siberia. Each glacial advance tied up large volumes of water and the sea levels dropped globally by around 100 m.
During the interglacials, the sea level rose again and coastal flooding was common during this time.
The most important sites for beetle fossils of the Miocene are situated in the warm temperate and to subtropical zones. Many recent genera and species already existed during the
Miocene, however, their distribution differed considerably from today's. One of the most important fossil sites for insects of the Pliocene is Willershausen
near Göttingen, Germany with excellently preserved beetle fossils of various families (longhorn beetles, weevils, ladybugs and others) as well as representatives of other orders
of insects40. In the Willershausen clay pit so far 35 genera from 18 beetle families have been recorded, of with six genera are extinct.
The Pleistocene beetle fauna is relatively well known, for instance in the scientific work of S. A. ELIAS41-43,
who used the composition of the beetle fauna to reconstruct climate conditions in the Rocky Mountains and on Beringia, the former land bridge between Asia and North America.

Fig. 14: Pterolophosoma otiliae, Dominican amber, Lower Miocene44. © Entomapeiron / Francesco Vitali, use with written permission.
|
3. Summary |
|
From an evolutionary point of view, the order of the beetles is matchless. Today, 166 families are known, compared to 67 from the Paleozoic and the Mesozoic.
19 of these families, including all from the Paleozoic became extinct45 (compare Fig. 15). Many recent families can be
traced back the relatives from the Cretaceous, partially even from the Jurassic or Triassic - contrary to many other families of the animal kingdom, which can
only be traced back to the Paleogene. In other words, the evolution of the beetles was more continuous and less rapid than of many other terrestrial animals.
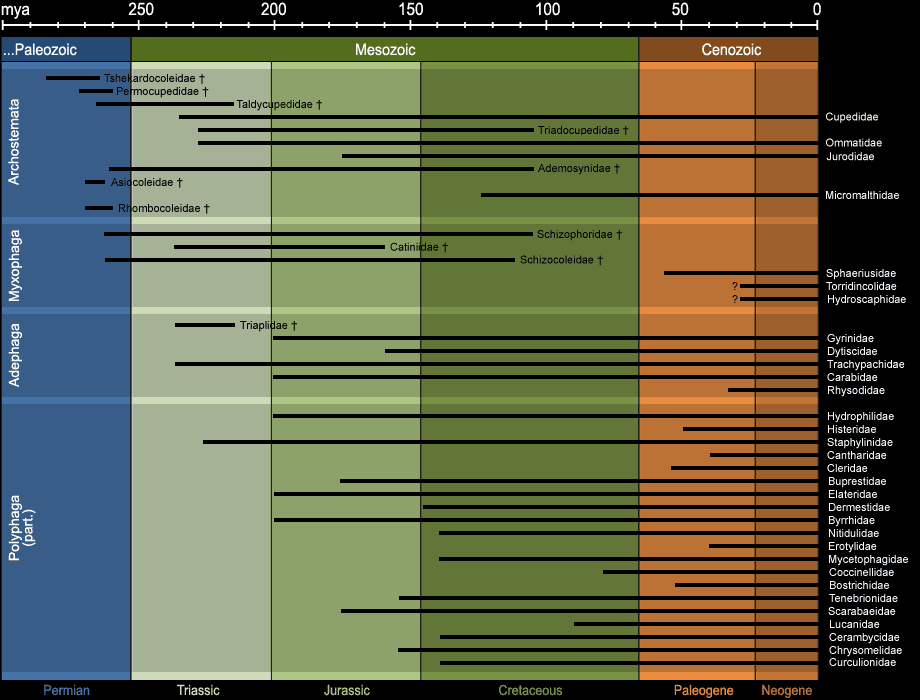
Fig. 15: Appearance of selected extinct and recent beetle families in Earth history. The figure has been compiled from the literature and is strongly simplified.
In 2007, A. P. VOGLER stated in his remarkable paper on the phylogeny of the beetles in Science2,
that "the success of beetles is explained neither by exceptional net diversification rates nor by a predominant role of herbivory and the Cretaceous rise of angiosperms.
Instead, the pre-Cretaceous origin of more than 100 present-day lineages suggests that beetle species richness is due to high survival of lineages and sustained
diversification in a variety of niches".
Maybe the famous (and probably apocryphal) quote of the geneticist and evolutionary biologist J. B. S. HALDANE (1892-1964) can answer the question,
why there are so many beetles. When asked by some theologians, what could be inferred about the mind of the Creator from the works of His Creation,
HALDANE answered: "An inordinate fondness of beetles".
|
Thanks |
|
It would have been impossible to make this page without the generous support with photographs by a number of scientists from paleontology and geology.
The support with photographs and maps by the following scientists is gratefully acknowledged:
- Ren Dong and Huali Chang, College of Life Science, Capital Normal University, Beijing, China
- Rafael G. Martins-Neto, Universidade Federal de Juiz de Fora, Juiz de Fora, Brazil
- Oscar F. Gallego, Universidad Nacional del Nordeste, Corrientes, Argentina
- Sonja Wedmann, Research Institute Senckenberg, Research Station Messel Pit, Messel, Germany
- Carsten Brauckmann and Elke Gröning, Institute for Geology and Paleontology, TU Clausthal, Germany
- Roy Beckemeyer, Museum Associate of the University of Kansas Natural History Museum, Lawrence, KS, USA
- Ron Blakey, Northern Arizona University, Department of Geology, Flagstaff, AZ, USA
- Francesco Vitali, Genoa, Italy
- Doug Lundberg, Amberica West, USA
For the review of the draft and helpful discussions I want to thank Irina Würtele and Klaas Reißmann.
|
Literature
- Die Insektentaphozönose von Niedermoschel (Asselian, unt. Perm; Deutschland)
T. Hörnschemeyer, H. Stapf, Terra Nostra. Schriften der Alfred-Wegener-Stiftung 99/8: 98.
- On the systematic position of the supposed Permian beetles, Tshecardocoleidae [sic], with a description of a new collection from Moravia
J. Kukalová, Sb. Geol. Ved. Rada. P. Paleontol. 1969, 11, 139-161.
- A new family of Coleoptera from the Permian of the Urals
B. B. Rohdendorf, Dokl. Akad. Nauk. SSSR 1944, 44, 252-253.
- Paleozoic beetles of Cupedidae from the European USSR
A. G. Ponomarenko, Paleontol. Zh. 1963, 1, 70-85.
- Permocoleus, New Genus, the First Permian Beetle (Coleoptera) from North America
S. H. Lubkin, M. S. Engel Ann. Entomol. Soc. Am. 2005, 98(1), 73-76. 
- A Second Specimen of Permocoleus (Coleoptera) from the Lower Permian Wellington Formation of Noble County, Oklahoma
R. J. Beckemeyer, M. S. Engel, Journal of the Kansas Entomological Society 2008, 81(1), 4-7.
- The First Beetles (Permosynidae, Coleoptera) from the Upper Tatarian of European Russia
A. G. Ponomarenko, Paleontological Journal 2003, 37(2), 170-173.
- New beetles (Insecta: Coleoptera) from the Late Permian of South Africa
A. G. Ponomarenko, M. B. Mostovski, African Invertebrates 2005, 46, 253-260.
- On Permian and Triassic Insect Faunas in Relation to Biogeography and the Permian-Triassic Crisis
D. E. Shcherbakov, Paleontological Journal 2008, 42(1), 15-31.
- Beetles (Insecta, Coleoptera) of the Late Permian and Early Triassic
A. G. Ponomarenko, Paleontological Journal 2004, 38 (Suppl. 2), S185-S196.
- New Triassic Beetles (Coleoptera) from Northern European Russia
A. G. Ponomarenko, Paleontological Journal 2008, 42(6), 600-606.
- Fossil insects from the Tithonian "Solnhofener Plattenkalke" in the Museum of Natural History, Vienna
A. G. Ponomarenko, Ann. Naturhist. Mus. Wien 1985, 87, 135-144.
- A New Genus of Elateriform Beetles (Coleoptera, Polyphaga) from the Middle-Late Jurassic of Karatau
E. V. Yan, Paleontological Journal 2009, 43 (1), 78-82.
- New Ommatids from the Late Jurassic of western Liaoning, China (Coleoptera: Archostemata)
J.-J. Tan, D. Ren, M. Liu, Insect Science 2005, 12, 207-216.
- New Cupedids from the Middle Jurassic of Inner Mongolia, China (Coleoptera: Archostemata)
J.-J. Tan, D. Ren, C. Shih, Annales Zoologici (Warszawa) 2006, 56(1), 1-6.
- Palaeogeography, Palaeoecology and Taphonomy of Jurassic-Cretaceous Cupedomorpha Faunas from China
J.-J. Tan, D. Ren, C. Shih, Mesozoic Terrestrial Ecosystems 2006, 130-136.
- New Beetles of the Family Cupedidae from the Mesozoic of Mongolia. Ommatini, Mesocupedini, Priacmini
A. G. Ponomarenko, Paleontological Journal 1997, 31(4), 389-399.
- Early Jurassic Insects from the Newark Supergroup, Northeastern United States
P. Huber, N. G. McDonald, P. E. Olsen in P. M. LeTourneau and P.E. Olsen (eds.), The Great Rift Valleys of Pangea in Eastern
North America, 2003, Volume 2: Sedimentology, Stratigraphy, and Paleontology, Columbia University Press, New York, 206-223.
- Insect diversity in the fossil record
C. C. Labandeira, J. J. Sepkoski, Science 1993, 261, 310-315.
- The fossil record of weevils and related beetle families (Coleoptera, Curculionoidea)
V. G. Gratshev, V. V. Zherikhin, Acta zoologica cracoviensia 2003, 46, 129-138.
- A new genus and two new species of fossil Elaterids from the Yixian Formation of Western Liaoning, China (Coleoptera: Elateridae)
H. Chang, F. Zhang, D. Ren, Zootaxa 2008, 1785, 54-62.
- New Fossil Elaterids (Coleoptera: Polyphaga: Elateridae) From the Jehol Biota in China
H. Chang, A. Kirejtshuk, D. Ren, Annals of the Entomological Society of America, 2010, 103(6), 866-874.
- A Survey of Mesozoic Buprestids (Coleoptera) from Eurasian deposits
A. V. Alexeev, Proceedings of the First International Palaeoentomological Conference, Moscow 1998.
- On Mesozoic Buprestids (Coleoptera: Buprestidae) from Russia, Kazakhstan, and Mongolia
A. V. Alexeev, Paleontological Journal 2000, 34(3), S323-S326.
- Jurassic and Lower Cretaceous Buprestidae (Coleoptera) from Eurasia
A. V. A. Orekhovo-Zuyevo, Paleontological Journal 1993, 27(1A), 9-34.
- The Crato Fossil Beds of Brazil - Window into an Ancient World
D. M. Martill, G. Bechly and R. F. Loveridge, Cambridge University Press 2007, ISBN 978-0-521-85867-0.
- New beetles (Insecta: Coleoptera) from the Lower Cretaceous of Spain
A. Ponomarenko, X. Martínez-Delclòs, Acta Geologica Hispanica 2000, 35(1-2), 47-52.
- New Jewel Beetles (Coleoptera: Buprestidae) from the Cretaceous of Russia, Kazakhstan, and Mongolia
Paleontological Journal 2009, 43(3), 277-281.
- The Role of Insects in Late Jurassic to Middle Cretaceous Ecosystems
C. C. Labandeira in S. G. Lucas, J. I. Kirkland and J. W. Estep (eds.), 1998, Lower and Middle Cretaceous
Terrestrial Ecosystems, New Mexico Museum of Natural History and Science Bulletin No. 14., 105-124.
- Dinosaurs, dung beetles, and conifers; participants in a Cretaceous food web
K. Chin, and B. D. Gill, Palaios 1996, 11(3), 280-285.
- Did dinosaurs have any relation with dung-beetles? (The origin of coprophagy)
A. Arillo, V. M. Ortuño, Journal of Natural History 2008, 42(19&20), 1405-1408.
- Timing the Radiations of Leaf Beetles: Hispines on Gingers from Latest Cretaceous to Recent
P. Wilf, C. C. Labandeira, W. J. Kress, C. L. Staines, D. M. Windsor, A. L. Allen, K. R. Johnson, Science 2000, 289, 291-294.
- A New Weevil-Beetle (Insecta, Coleoptera, Nemonychidae) from the Lower Cretaceous of Spain
V. V. Zherikhin and V. G. Gratshev, Paleontological Journal 2003, 37(4), 407-408.
- Fossil Curculionoid Beetles (Coleoptera, Curculionoidea) from the Lower Cretaceous of Northeastern Brazil
V. V. Zherikhin and V. G. Gratshev, Paleontological Journal 2004, 38(5), 528-537.
- New fossil dermestid beetles (Coleoptera: Dermestidae) from the Baltic amber - II
J. Háva, J. Prokop, M. Kadej, Studies and reports of District Museum Prague-East Taxonomical Series 2, 2006, 1-2, 65-68.
- New fossil dermestid beetles (Coleoptera: Dermestidae) from the Baltic amber - III
J. Háva, J. Prokop, A. Herrmann, Acta Soc. Zool. Bohem. 2007, 71, 151-157.
- Osoriinae (Coleoptera: Staphylinidae) from Dominican Amber
U. Irmler, Stuttgarter Beitr. Naturk. Ser. B 2003, 342, 1-16.
- New Amber Deposit provides evidence of early Paleogene extinctions, paleoclimates, and past distibutions
G. Poinar Jr., B. Archibald, A. Brown, The Canadian Entomologist 1999, 131, 171-177.
- Fossile Prachtkäfer (Coleoptera: Buprestidae: Buprestinae) aus dem Mitteleozän der Grube Messel bei Darmstadt, Teil 1
T. Hörnschemeyer, S. Wedmann, Cour. Forsch.-Inst. Senckenberg 1994, 170, 85-136.
- Dritter Beitrag über Käfer (Coleoptera) aus dem Jungtertiär von Willershausen, Bl. Northeim 4226
E. Gersdorf, Geol. Jahrb. A 1976, 36, 103-145.
- Late Pleistocene and Holocene Seasonal Temperatures Reconstructed from Fossil Beetle Assemblages in the Rocky Mountains
S. A. Elias, Quaternary Research 1996, 46 (3), 311-318.
- Late Pleistocene Climates of Beringia, Based on Analysis of Fossil Beetles
S. A. Elias, Quaternary Research 2000, 53 (2), 229-235.
- Climatic Tolerances and Zoogeography of the Late Pleistocene Beetle Fauna of Beringia
S. A. Elias, Géographie physique et Quaternaire 2000, 54 (2), 143-155.
- A new cerambycid from Dominican amber and remarks on the fossil Plectromerus-species (Coleoptera Cerambycidae)
F. Vitali, Entomapeiron Paleoentomology 1(1): 1-12
- The geological history of beetles
A. G. Ponomarenko in J. Pakaluk, S. A. Ślipiński (eds.), 1995, Biology, phylogeny, and classification of Coleoptera:
papers celebrating the 80th birthday of Roy A. Crowson, 155-171. Museum i Instytut Zoologii PAN, Warszawa.
|
Further Reading
- Evolution and Classification of Beetles
J. F. Lawrence, A. F. Newton Jr., Ann. Rev. Ecol. Syst. 1982, 13, 261-290.
- A Comprehensive Phylogeny of Beetles Reveals the Evolutionary Origins of a Superradiation
A. P. Vogler et al., Science 2007, 318, 1913-1916.
- "Inordinate Fondness" Explained: Why Are There So Many Beetles?
B. D. Farrell, Science 1998, 281, 555-559.
- Fossil insects in Gondwana - localities and palaeodiversity trends
T. Schlüter, Acta zoologica cracoviensia 2003, 46, 345-371.
- Ecological evolution of beetles (Insecta: Coleoptera)
A. G. Ponomarenko, Acta zoologica cracoviensia 2003, 46, 319-328.
- Evolutionary Rates in the Adaptive Radiation of Beetles on Plants
B. D. Farrell, A. S. Sequeira, Evolution 2004, 58(9), 1984-2001.
- The Evolution of Agriculture in Beetles (Curculionidae: Scolytinae and Platypodinae)
B. D. Farrell et al., Evolution 2001, 55(10), 2011-2027.
|

 Fig. 12: The jewel beetle Psiloptera weigelti, Messel pit, Germany, 47 mya. © Research institute Senckenberg, use with written permission.
Fig. 12: The jewel beetle Psiloptera weigelti, Messel pit, Germany, 47 mya. © Research institute Senckenberg, use with written permission.











