Entwicklungsgeschichte der Käfer |
Text © Christoph Benisch, 2010
|
|
|
1. Einführung |
|
Die Käfer sind entwicklungsgeschichtlich gesehen eine alte Ordnung (s. Abb. 1a). Die ältesten fossilen Funde stammen aus dem Perm, lange bevor die Dinosaurier
die Welt beherrschten. Seither haben sich die Käfer zur artenreichsten Ordnung im Tierreich überhaupt entwickelt. Weltweit sind
über 350.000 Arten beschrieben. Das bedeutet, dass Käfer heute 40% aller wissenschaftlich beschriebenen Insekten und 25% aller bisher beschriebenen Tierarten
stellen. Auch heute noch wird jährlich eine große Zahl Arten neu entdeckt und Schätzungen gehen von bis zu 8 Mio. Käferarten weltweit aus.
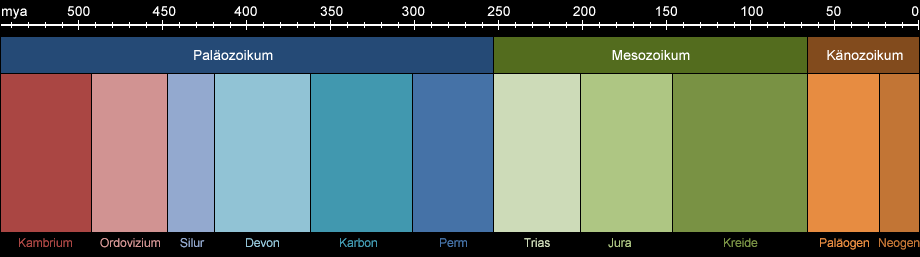
Abb. 1a: Das Erdzeitalter des Phanerozoikums. Es umfasst die Ären vom Paläozoikum bis heute. Anklicken, um die Entstehung ausgewählter Tiergruppen anzuzeigen.
Die globalen klimatischen Verhältnisse änderten sich im Verlauf der Erdgeschichte mehrfach. Mittels Sedimentanalyse lassen sie sich
bis vor etwa 2,3 Milliarden Jahren rekonstruieren. Während des Phanerozoikums gab es mehrfach Eiszeiten, die durch vollständige Vereisung
der Polkappen gekennzeichnet sind. Auch heute leben wir erdgeschichtlich gesehen in einer Eiszeit.
Dies ist keineswegs der "Normalzustand" der Erde. Vielmehr herrschten in den letzten 550 Mio. Jahren in etwa 80% der Zeit akryogene (nicht eisbildende) Warmzeiten vor, in
denen die Polkappen vollkommen eisfrei waren. Abb. 1b zeigt den durchschnittlichen Temperaturverlauf in vereinfachter Form.
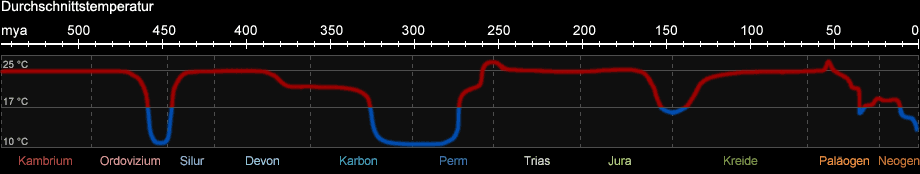
Abb. 1b: Klimatische Verhältnisse seit dem Paläozoikum (nach C. R. Scotese, Paleomap, 2008)
Das silur-ordovizische Eiszeitalter begann etwa vor 450 Mio. Jahren und war auf das heutige Nordafrika und Südamerika beschränkt. Aufgrund
der starken Abkühlung der Meere war es von einem Massenaussterben von Arten begleitet (s. Abb. 1c). Es folgte eine Warmzeit, die vom oberen Silur bis zum mittleren
Karbon anhielt. Gegen Ende des Karbons setzte die permokarbonische Eiszeit ein, in der die Vergletscherung im Wesentlichen auf die südliche Hemisphäre beschränkt war und die
daher auch als "Gondwana-Vereisung" bezeichnet wird. Vor ca. 2,6 Mio. Jahren begann die letzte und bis heute andauernde Eiszeit, die auch
als "Quartäres Eiszeitalter" bezeichnet wird. Sie ist gekennzeichnet durch den schnellen Wechsel sehr kalter Abschnitte (Glaziale) und dazwischenliegender
Warmzeiten (Interglaziale).
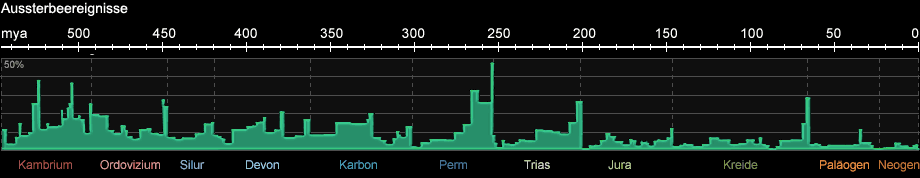
Abb. 1c: Aussterbeereignisse während des Phanerozoikums (nach R. A. Rohde und R. A. Muller, 2005)
|
2.0 Perm (299-250 mya) |
|
Während des Perm waren die Landmassen zu einem einzigen Superkontinent (Pangäa) vereinigt (vgl. Abb. 2). Dieser bildete sich durch den Zusammenstoß des bereits
im Karbon vereinigten Großkontinents Gondwana (das heutige Südamerika, Afrika, Antarktis, Australien, Madagaskar und Indien) und Laurussia
(das heutige Europa und Nordamerika) mit Sibiria (das heutige mittelsibirische Bergland). Durch die Kollision wurde der Ural aufgefaltet. Der Superkontinent war von einem
einzigen Meer umgeben, dem sogenannten Panthalassa. Das in der östlichen Ausbuchtung von Pangäa befindliche Meer wird Tethys genannt. Wesentliche Teile des heutigen
Europas waren von einem salzreichen Binnenmeer, dem Zechsteinmeer, bedeckt, dessen Becken im Verlauf des Perm mehrfach austrocknete und sich wieder auffüllte.
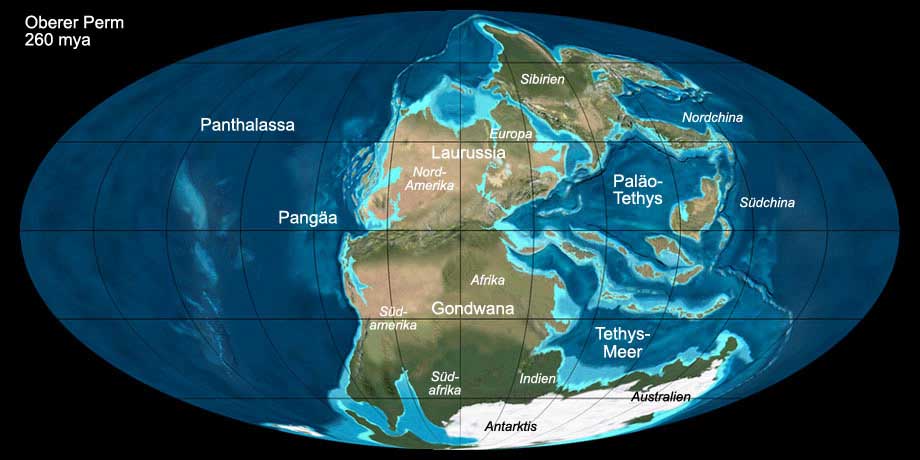
Abb. 2: Die Welt während des oberen Perm (260 mya). Anklicken, um Klima und Beschriftung ein-/auszublenden. © Ron Blakey, Verwendung mit schriftlicher Genehmigung.
Das Klima zu Beginn des Perm war zunächst durch die permokarbonische Vereisung gekennzeichnet. Die Polregionen waren durch massive Eisschichten
bedeckt, wesentliche Teile von Gondwana waren ebenfalls vergletschert. In den tropischen Regionen breiteten sich Sumpfwälder aus. Während des mittleren
Perm wurde das Klima wärmer und trockener und die Gletscher wichen zurück. Das Landesinnere war durch kontinentales Klima mit starken Temperaturschwankungen
und Regen-/Trockenzeiten gekennzeichnet.
| System |
Serie |
Stufe |
Alter (mya) |
| Perm |
Lopingium |
Changhsingium |
251,0 - 253,8 |
| Wuchiapingium |
253,8 - 260,4 |
| Guadalupium |
Capitanium |
260,4 - 265,8 |
| Wordium |
265,8 - 268,0 |
| Roadium |
268,0 - 270,6 |
| Cisuralium |
Kungurium |
270,6 - 275,6 |
| Artinskium |
275,6 - 284,4 |
| Sakmarium |
284,4 - 294,6 |
| Asselium |
294,6 - 299,0 |
Die frühesten fossilen Käferfunde stammen aus dem unteren Perm (Asselium-Stufe) und sind ca. 300 Mio. Jahre alt.
Sie finden sich z.B. in der Niedermoschelbank im unteren Rotliegenden in der Nähe von Mainz1. Weitere bekannte Funde stammen aus Obora,
Tschechien2 (Artinskium-Stufe) und Tshekarda im Ural, Russland3,4 (Kungurium-Stufe). Interessanterweise wurden
aus der Artinskium-Stufe im heutigen Nordamerika im Gegensatz zu Europa bislang kaum fossile Käfer nachgewiesen, obwohl beide Kontinente zu dem Großkontinent
Laurussia vereinigt waren. Die beiden ersten Nachweise aus dem heutigen Nordamerika überhaupt wurden in der Wellington-Formation in Oklahoma erbracht und
2005 bzw. 2008 publiziert5,6.
In den oberen Stufen des mittleren Perm sind die Käfer in Europa und dem europäischen Teil Asiens eher selten nachgewiesen worden. PONOMARENKO
publizierte 2003 den ersten fossilen Fund aus der Tatarium-Stufe (das regionale Äquivalent der Capitanium-Stufe)7.
Im oberen Perm tauchten Käfer dann in allen Erdteilen auf, vor allem Laurussia und Sibiria, später auch in Gondwana.
Fossile Funde aus dem oberen Perm wurden in Südafrika8, Südamerika, Australien und Osteuropa gemacht. Bei der überwiegenden Mehrzahl
dieser Funde handelt es sich um (meist isolierte) Flügeldecken. Diese hatten sich während des Perm durch Umbildung des vorderen häutigen
Flügelpaars zu hartschaligen Flügeldecken entwickelt. Diese Entwicklung war am Ende des Perm abgeschlossen.
Abb. 3: Flügeldecke von Permocoleus wellingtonensis, Wellington-Formation, Oklahoma, USA (Unterer Perm, Artinskium-Stufe). Anklicken zum Vergrößern.
© Roy Beckemeyer, Verwendung mit schriftlicher Genehmigung.
Am Ende des Perm kam es zum größten Massenaussterben in der Erdgeschichte (das sog. PT-Ereignis): 95% aller meeresbewohnenden Arten und 66% aller
an Land lebenden Arten starben aus. Auch 30% aller Insektenarten waren betroffen - es war das bis heute einzige Massenaussterben von Insekten in der
Erdgeschichte. Die Gründe für das Artensterben sind bis heute nicht geklärt, als Ursachen werden Vulkanismus, ein Meteoriteneinschlag
oder ein kosmischer Gammablitz diskutiert.
|
2.1 Trias (251-199 mya) |
|
Bereits im oberen Perm begann Pangäa auseinanderzubrechen, es bildeten sich Riftsysteme zwischen dem späteren Nordamerika und Afrika.
Während der Trias erweiterte sich die Tethys Richtung Westen (Neotethys). In der Obertrias entstanden zwischen dem späteren Nordamerika und Europa
weitere Riftsysteme und es kam zur marinen Ingression.
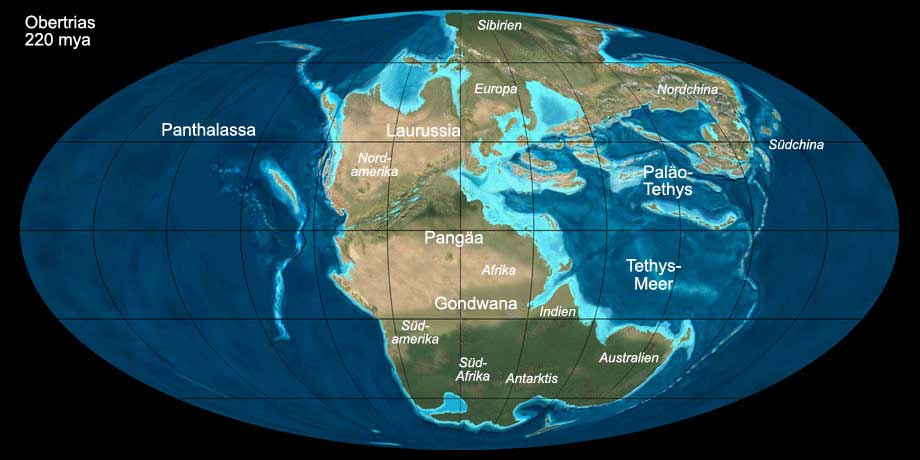
Abb. 4: Die Welt während der Obertrias (220 mya). Anklicken, um Klima und Beschriftung ein-/auszublenden. © Ron Blakey, Verwendung mit schriftlicher Genehmigung.
Das Klima während der gesamten Trias war warm bis heiß und überwiegend trocken, im Landesinneren sogar ausgesprochen arid und stark kontinental geprägt
(sehr heiße Sommer und kalte Winter). Der Meeresspiegel war während der Trias niedrig und trug so zum warmen Klima bei. Die Pole waren eisfrei.
| System |
Serie |
Stufe |
Alter (mya) |
| Trias |
Obertrias |
Rhaetium |
199,6 - 203,6 |
| Norium |
203,6 - 216,5 |
| Karnium |
216,5 - 228,7 |
| Mitteltrias |
Ladinium |
228,7 - 237,0 |
| Anisium |
237,0 - 245,9 |
| Untertrias |
Olenekium |
245,9 - 249,5 |
| Indusium |
249,5 - 251,0 |
Aus der gesamten Untertrias sind als Folge des Massenaussterbens an der Grenze zwischen Perm und Trias nur wenige fossile Insekten,
einschließlich Käfer nachgewiesen9. Eine Ausnahme stellen Funde in Osteuropa dar: In der Fundstätte Babiy Kamen10
im Kusnezkbecken (Indusium- und Olenekium-Stufe) wurden zahlreiche fossile Käfer nachgewiesen, auch vollständig erhaltene Exemplare aus den
Unterordnungen Archostemata (Ademosynidae, Schizocoleidae), Adephaga (Triaplidae, Trachypachidae) und Polyphaga
(Hydrophilidae, Byrrhidae, Elateroidea).
Käfer der Familien Cupedidae und Schizophoroidae sind hier dagegen nicht vertreten, während sie an anderen Fundstätten aus der Untertrias dominieren.
Weitere Funde sind aus Khey-Yaga11 im Korotaikha-Becken (Olenekium- und Anisium-Stufe) bekannt.
Ab der Mitteltrias erscheinen mycetophage Cupedidae. In den Stufen der Obertrias wurden Vertreter der algenfressenden Triaplidae und Hydrophilidae nachgewiesen,
ebenso räuberisch lebende Wasserkäfer. Mit den Obrienidae erscheinen auch erste primitive Rüsselkäfer-Arten, sowie die ersten Vertreter
der Kurzflügler (Staphylinidae), die sich im Körperbau nicht wesentlich von den heutigen Arten unterscheiden.
 Abb. 5: Flügeldecke von Argentinosyne duraznoensis (Schizocoleidae), Potrerillos Formation, Mendoza Province, Argentinien (Frühe Obertrias, Karnium-Stufe).
© Oscar F. Gallego, Verwendung mit schriftlicher Genehmigung.
Abb. 5: Flügeldecke von Argentinosyne duraznoensis (Schizocoleidae), Potrerillos Formation, Mendoza Province, Argentinien (Frühe Obertrias, Karnium-Stufe).
© Oscar F. Gallego, Verwendung mit schriftlicher Genehmigung.
|
2.2 Jura (199-145 mya) |
|
Während des Juras zerfiel der Superkontinent Pangäa, der bereits während der Trias begonnen hatte auseinanderzubrechen, weiter in den nördlichen Teil Laurasia
und den südlichen Teil Gondwana. Der Nordatlantik zwischen Europa und Nordamerika war noch sehr schmal, der Südatlantik öffnete sich erst gegen Ende des Juras.
In Nordamerika begannen sich die Rocky Mountains aufzufalten. Die Antarktis wurde durch einen Graben von Afrika und Südamerika getrennt und das heutige
Indien begann Richtung Norden zu driften. Der Meeresstand war höher als heute und weite Teile Europas und Nordamerikas waren von einem flachen tropischen
Meer von großer Artenvielfalt bedeckt.
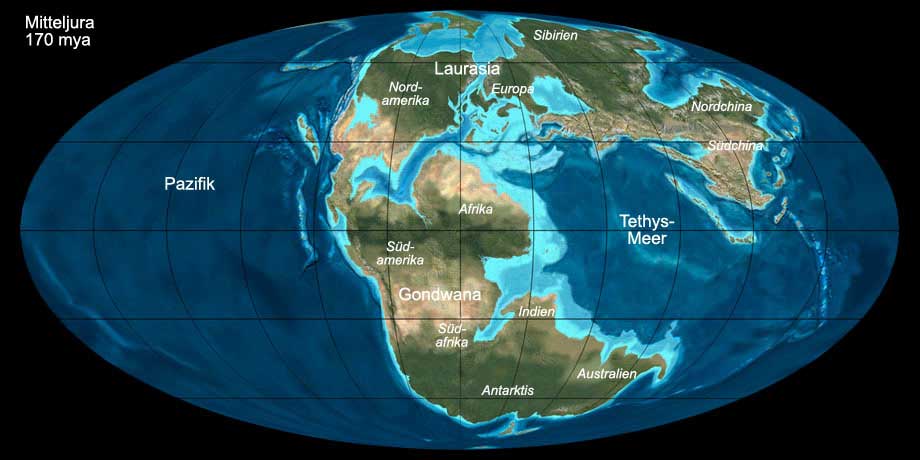
Abb. 6: Die Welt während des Mitteljuras (170 mya). Anklicken, um Klima und Beschriftung ein-/auszublenden. © Ron Blakey, Verwendung mit schriftlicher Genehmigung.
Das globale Klima während des Juras war warm und feucht. Wie in der Trias befanden sich während des Juras in der Nähe der Pole keine größeren Landmassen, dementsprechend waren
keine Inlandeisschilde vorhanden. Obgleich manche Bereiche des heutigen Nord- und Südamerika und Afrika arid blieben, waren weite Teile der kontinentalen Landmasse üppig bewachsen.
Die nördliche (laurasische) und südliche (gondwanische) Fauna war anfänglich noch deutlich verschieden. Während des Juras bekam sie jedoch einen interkontinentalen
Charakter und zahlreiche Arten verbreiteten sich weltweit.
| System |
Serie |
Stufe |
Alter (mya) |
| Jura |
Oberjura |
Tithonium |
145,4 - 150,8 |
| Kimmeridgium |
150,8 - 155,7 |
| Oxfordium |
155,7 - 161,2 |
| Mitteljura |
Callovium |
161,2 - 164,7 |
| Bathonium |
164,7 - 167,7 |
| Bajocium |
167,7 - 171,6 |
| Aalenium |
171,6 - 175,6 |
| Unterjura |
Toarcium |
175,6 - 183,0 |
| Pliensbachium |
183,0 - 189,6 |
| Sinemurium |
189,6 - 196,5 |
| Hettangium |
196,5 - 199,6 |
Aus dem Jura sind weltweit über 150 bedeutende Fundstätten fossiler Käfer bekannt, die Mehrzahl liegt in (Ost-) Europa und Nordasien.
In Nordamerika und vor allem in Südamerika und Afrika ist die Anzahl der Lagerstätten aus dem Jura geringer und sie sind bislang wenig untersucht.
Herausragende Lagerstätten sind Solnhofen12 (Tithonium-Stufe) in Oberbayern, Deutschland, Karatau13 in Südkasachstan (Oxfordium- oder
Kimmeridgium-Stufe), die Yixian-Formation in Liaoning14, Nordchina (Tithonium-Berriasium-Stufe,
enthält die Jehol-Biota) sowie die Jiulongshan-Formation15,16 (Aalenium- oder Bajocium-Stufe, enthält die Yanliao-Biota)
und weitere Lagerstätten17 in der Mongolei. In Nordamerika sind die wenigen bekannten Fundstätten von Insekten aus dem Jura in den Muschelkalk-Ablagerungen im Hartford-, Deerfield-
und Newark-Becken18 (Hettangium-Sinemurium-Stufe) bekannt.
Die Käfer aus dem Unterjura sind den aus der oberen Trias bekannten Arten ähnlich. Die xylophagen Cupedidae traten nur in den warmen Zonen auf.
Zum Oberjura hin nahm der Anteil der Cupedidae ab und gleichzeitig nahm die Diversität der frühen phytophagen Arten zu. Die meisten rezenten phytophagen
Käferarten leben an Bedecktsamern (Angiospermae). Es wird angenommen, dass die Zunahme der Diversität bei den Bedecktsamern auch die Diversität der
phytophagen Arten beeinflusste, die sich während des Mitteljuras verdoppelte. Allerdings wird diese Hypothese in jüngerer Zeit wieder angezweifelt,
da die Zunahme der Anzahl der Käferfamilien während der Kreide nicht mit der Vergrößerung der Anzahl der Bedecktsamer-Arten korreliert19.
Im Oberjura kamen bereits zahlreiche ursprüngliche Rüsselkäfer20 (Curculionoidea) und Schnellkäfer21,22
(Elateroidea) vor. Auch erste Prachtkäfer23-25 (Buprestidae) sind nachweisbar, allerdings blieben sie bis zur Kreide relativ selten.
Abb. 7: 1a,b: Paradesmatus baiae; 6a,b: Paradesmatus ponomarenkoi; 9: Protagrypnus robustus; Jiulongshan Formation, Ningcheng-Provinz, Innere Mongolei, China (Mitteljura, Aalenium- oder Bajocium-Stufe). Anklicken zum Vergrößern.
© Ren Dong und Huali Chang, Verwendung mit schriftlicher Genehmigung.
|
2.3 Kreide (145-65 mya) |
|
Die Kreide ist die jüngste Periode des Mesozoikums und währte 80 Mio. Jahre (von 145 bis 65 mya).
Während der Kreide brach der frühere Superkontinent Pangäa vollends auseinander. Die während der Unterkreide noch zusammenhängenden Landmassen von Afrika und Südamerika
trennten sich und öffneten den Südatlantik. Ab dem Turonium bestand eine Verbindung zum Nordatlantik.
Australien und Indien begannen sich von der Antarktis zu trennen und nach Norden zu driften. Madagaskar und Indien blieben noch verbunden. Weite Teile
Nordamerikas und Europas waren von einem flachen Meer bedeckt, welches sich erst am Ende der Kreide zurückzog.
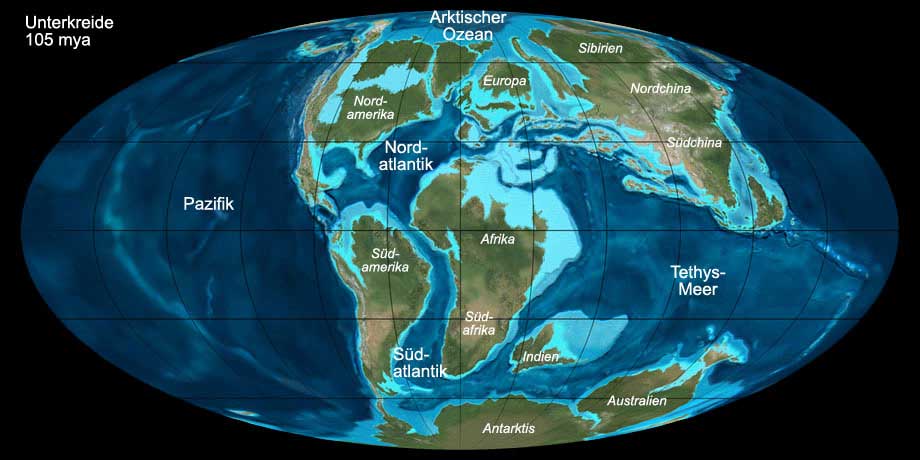
Abb. 8: Die Welt während der späten Unterkreide (105 mya). Anklicken, um Klima und Beschriftung ein-/auszublenden. © Ron Blakey, Verwendung mit schriftlicher Genehmigung.
Zu Beginn der Kreide, im Berriasium, setzte sich die Abkühlung fort, die bereits im Oberjura begonnen hatte. Vermutlich schneite es in höheren Lagen,
zur Vergletscherung außerhalb der Hochlagen kam es indessen nicht. Am Ende des Berriasiums stiegen die Temperaturen wieder an und blieben bis zum
Ende der Kreide relativ konstant. Durch Vulkanismus stieg die CO2-Konzentration in der Atmosphäre an und trug zur Erwärmung bei.
Das Klima an den Polen war kühl gemäßigt (Dinosaurier kamen bis zum 15. Breitengrad um den damaligen Südpol vor). Der sehr flache Temperaturgradient
von den Tropen zu den Polen verursachte global nur schwache Winde, die Ozeane waren weniger bewegt und Sauerstoffmangel kam häufig vor, wie durch
Tonschiefer-Sedimente belegt ist.
| System |
Serie |
Stufe |
Alter (mya) |
| Kreide |
Oberkreide |
Maastrichtium |
65,5 - 70,6 |
| Campanium |
70,6 - 83,5 |
| Santonium |
83,5 - 85,8 |
| Coniacium |
85,8 - 88,6 |
| Turonium |
88,6 - 93,5 |
| Cenomanium |
93,5 - 99,6 |
| Unterkreide |
Albium |
99,6 - 112,0 |
| Aptium |
112,0 - 125,0 |
| Barremium |
125,0 - 130,0 |
| Hauterivium |
130,0 - 133,9 |
| Valanginium |
133,9 - 140,2 |
| Berriasium |
140,2 - 145,5 |
Weltweit sind zahlreiche bedeutende Lagerstätten fossiler Käfer aus der Kreide bekannt. Die meisten liegen in Europa und Asien und gehörten
damals zur klimatisch gemäßigten Zone. Einige der im Abschnitt Jura genannten Lagerstätten aus dem Oberjura geben auch Aufschluss über die Käferfauna
der Unterkreide, z.B. die Yixian-Formation in Liaoning14, Nordchina (Tithonium-Berriasium-Stufe). Weitere wichtige
Lagerstätten der Unterkreide sind die Crato-Schicht26 im Araripe-Becken im Bundesstaat Ceará, Nordbrasilien
(Aptium-Albium-Stufe) sowie die darüber liegende Santana-Schicht (Cenomanium-Stufe). Letztere lag in der Nähe des Paläoäquators.
In Spanien sind die Fundstätten bei Montsec (Hauterivium-Barremium-Stufe) und Las Hoyas27 (Barremium-Stufe) bedeutend,
in Australien die Koonwarra-Lagerstätte (Aptium-Stufe) der Korumburra-Gruppe, South Gippsland, Victoria.
Wichtige Lagerstätten aus der Oberkreide sind Kzyl-Dzhar (Turonium-Stufe) in Südkasachstan und Arkagala (Turonium-Stufe) in Russland.
Während der Kreide nahm die Artenvielfalt der Cupedidae und Archostemata deutlich ab. Räuberisch lebende Laufkäfer (Carabidae) und Kurzflügler
(Staphylinidae) zeigten eine ungleichmäßige Verbreitung. Während die Carabidae in den warmen Gebieten häufiger waren, kamen Staphylinidae eher
in den gemäßigten Klimazonen vor. Ebenfalls räuberisch lebten Cleroidea und Cucujoidea, die ihrer Beute unter der Rinde von Bäumen nachstellten.
Schnellkäfer (Elateridae) kamen in den warmen Gebieten häufig vor. Zusammen mit den Prachtkäfern (Buprestidae)28,
deren Artenvielfalt während der Kreide beträchtlich zunahm, waren sie an der biologischen Verwertung von Holz beteiligt29.
Bockkäfer (Cerambycidae) waren noch sehr selten und ihre Artenvielfalt nahm erst gegen Ende der Oberkreide zu.
 Abb. 9: Links: Zeichnung von Cratomacer immersus (Nemonychidae), Santana Formation, Crato Member, Ceará, Nordbrasilien (Aptium-Albium-Stufe),
nachgezeichnet nach V. V. Zherikhin und V. G. Gratshev, Paleontological Journal, 2004, 38(5), 530. Rechts: Alexcarabus megagnathus (Carabidae),
Santana Formation, Crato Member (s.o.), © Rafael G. Martins-Neto, Verwendung mit schriftlicher Genehmigung.
Abb. 9: Links: Zeichnung von Cratomacer immersus (Nemonychidae), Santana Formation, Crato Member, Ceará, Nordbrasilien (Aptium-Albium-Stufe),
nachgezeichnet nach V. V. Zherikhin und V. G. Gratshev, Paleontological Journal, 2004, 38(5), 530. Rechts: Alexcarabus megagnathus (Carabidae),
Santana Formation, Crato Member (s.o.), © Rafael G. Martins-Neto, Verwendung mit schriftlicher Genehmigung.
Die ersten Blatthornkäfer (Scarabaeidae) erschienen im Oberjura. Sie waren noch nicht coprophag, sondern ernährten sich wahrscheinlich von
verpilztem Holz. Dungkäfer sind erstmals in der Oberkreide (Campanium-Stufe) nachgewiesen worden30, sie lebten wahrscheinlich
an den Exkrementen pflanzenfressender Dinosaurier. Diskutiert wird, ob die Coprophagie der Käfer bereits bei ihrer Entwicklung in der Oberkreide an Säugetierexkremente
gebunden war31.
Unter den Schwimmkäfern finden sich die ersten Arten, bei denen sowohl die Larven als auch die Imagines an die aquatische Lebensweise angepaßt waren. Taumelkäfer
(Gyrinidae) waren mäßig artenreich, frühe Dytiscidae dagegen artenärmer. Am weitesten verbreitet waren die Coptoclavidae, die sich von wasserbewohnenden Fliegenlarven
ernährten.
Fraßspuren herbivorer Blattkäfer (Chrysomelidae) sind erstmals aus der Oberkreide nachgewiesen32. In der zweiten Hälfte
der Oberkreide verbreiteten sich die Donaciinae als Bewohner von Wasserpflanzen. Rüsselkäfer (Curculionidae) erschienen gegen Ende der Unterkreide.
Die Funde fossiler Käfer stammen vor allem aus Asien, vereinzelt auch aus Europa33 und Brasilien34.
Am Ende der Kreide vor 65 Mio. Jahren, der sog. KT-Grenze (Kreide-Tertiär, nach der früheren Bezeichnung des Paläogen) kam es zu einem großen Massenaussterben, das insbesondere
die Ära der Dinosaurier beendete. Man nimmt an, dass ein oder mehrere Meteoriteneinschläge und erhöhte Vulkanaktivität innerhalb eines Zeitraumes von etwa 1 Mio. Jahren
gravierende Umweltveränderungen verursacht haben. Die Theorie des Meteoriteneinschlags wird durch die Iridium- und Chrom-Anomalie in den Gesteinen an der KT-Grenze gestützt.
Käferarten scheinen von dem Massenaussterben wenig betroffen gewesen zu sein. So finden sich beispielsweise im östlichen Russland Gesteinsschichten auf
beiden Seiten der KT-Grenze, die eine vergleichbare Diversität aufweisen.
|
2.4 Paläogen (65-23 mya) |
|
Das Paläogen ist die älteste geochronologische Periode des Känozoikums und währte von 65 bis 23 mya. Während dieser Zeit nahm die Erdoberfläche
ihr heutiges Aussehen an. Die Fragmente Gondwanas (Südamerika, Afrika, Indien und Australien) begannen nordwärts zu driften. Der Zusammenprall
Indiens mit der eurasischen Landmasse führte zur Auffaltung des Himalaya, in ähnlicher Weise wurden die Alpen in Mitteleuropa durch die
Kollision der afrikanischen Landmasse mit Europa aufgefaltet. Südamerika driftete weniger weit nach Norden und der "Aufprall" wurde durch ein
System kleinerer Platten in der Karibik abgefedert, so dass die Auffaltung eines Gebirges unterblieb. Eine Landbrücke zwischen Nord- und Südamerika
bestand während des Paläogens noch nicht. Der Atlantik öffnete sich in Ost-West-Richtung während des Paläogens weiter. Im Norden brach die letzte
Landbrücke zwischen Nordamerika und Europa während des Eozän.
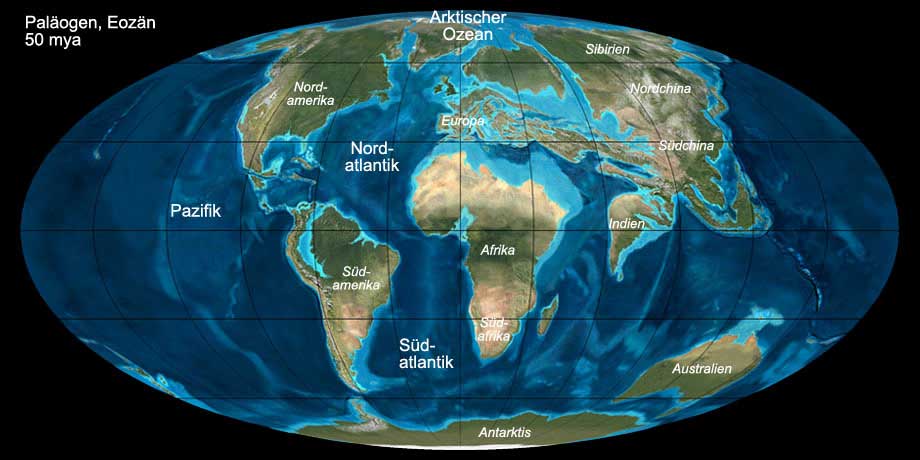
Abb. 10: Die Welt während des Paläogens (Eozän, 50 mya). Anklicken, um Klima und Beschriftung ein-/auszublenden. © Ron Blakey, Verwendung mit schriftlicher Genehmigung.
| System |
Serie |
Stufe |
Alter (mya) |
| Paläogen |
Oligozän |
Chattium |
28,4 - 23,0 |
| Rupelium |
33,9 - 28,4 |
| Eozän |
Priabonium |
37,2 - 33,9 |
| Bartonium |
40,4 - 37,2 |
| Lutetium |
48,6 - 40,4 |
| Ypresium |
55,8 - 48,6 |
| Paläozän |
Thanetium |
58,7 - 55,8 |
| Seelandium |
61,1 - 58,7 |
| Danium |
65,5 - 61,1 |
Das Klima während des Paläogens war warm und tropisch wie die meiste Zeit während des Mesozoikums. Zu Beginn (Paläozän) war das Klima zunächst noch
trockener und kühler als während der vorangegangenen Kreide, stieg aber zum Eozän stark an, so dass sich die subtropische Vegetationszone bis
nach Grönland und Patagonien erstreckte. Das Klima der Pole war kühl-gemäßigt, in Europa, Nordamerika, Australien und dem südlichen Teil Südamerikas dagegen warm-gemäßigt.
Am Äquator herrschte tropisches Klima, nördlich und südlich davon befanden sich heiße und trockene Zonen. Ab dem Oligozän begann die globale Abkühlung.
In der Antarktis bildete sich eine Eisdecke, wodurch der Meeresspiegel sank. Von einer Erwärmungsphase während des späten Oligozäns unterbrochen
setzte sich die Abkühlung fort und mündete schließlich in die Eiszeit des Pleistozäns.
Aus dem Paläogen sind zahlreiche fossile Funde von Käfern bekannt. Dabei ist die Käferfauna des Paläozäns (61-56 mya) vergleichsweise wenig untersucht.
Demgegenüber ist der Kenntnisstand der eozänen Käferfauna sehr hoch. Hierfür gibt es einen bestimmten Grund, nämlich das Auftreten von fossilen Insekten in
Bernstein und Tonschieferablagerungen. Bei Bernstein handelt es sich um fossile Baumharze, also um fossile organische Verbindungen und nicht um Minerale, wie es
der Name Bernstein suggeriert. Man unterscheidet Bernstein nach Ursprungsort, Alter und Art der produzierenden Pflanze. Für die Erforschung der eozänen
und oligozänen Käferfauna sind vor allem der baltische35,36 und der dominikanische Bernstein37
bedeutend.
 |
 |
 |
|
Abb. 11: Links: Sucinoptinus sucini, Baltischer Bernstein, oberes Eozän, © Francesco Vitali. Mitte: Curculionidae sp., Dominikanischer Bernstein, spätes Oligozän,
© Amberica West, USA. Rechts: Platypodidae sp., Dominikanischer Bernstein, spätes Oligozän, © Amberica West, USA. Verwendung mit schriftlicher Genehmigung.
|
 Abb. 12: Der Prachtkäfer Psiloptera weigelti, Grube Messel, Deutschland, 47 mya. © Forschungsinstitut Senckenberg, Verwendung mit schriftlicher Genehmigung.
Abb. 12: Der Prachtkäfer Psiloptera weigelti, Grube Messel, Deutschland, 47 mya. © Forschungsinstitut Senckenberg, Verwendung mit schriftlicher Genehmigung.
Baltischer Bernstein (Succinit) wird vor allem an den Küsten von Nord-
und Ostsee gefunden und stammt aus dem Eozän (überwiegend Lutetium, 50-40 mya) und wurde nach heutiger Auffassung von einer Verwandten der rezenten Goldlärche
(Pseudolarix) gebildet. Der dominikanische Bernstein stammt aus der Zeit vom Eozän bis zum unteren Miozän, das Harz wurde von Hymenaea protera gebildet. Der
dominikanische Bernstein zeichnet sich durch seine Klarheit und seinen Reichtum an Inklusen aus. Weitere interessante fossile Käferfunde stammen aus British Columbia,
Kanada38. Generell werden in Bernstein vor allem Vertreter der Familien Elateridae, Anobiidae, Scirtidae und Staphylinidae nachgewiesen.
Eine weitere Quelle fossiler Käfer sind Ölschieferablagerungen. Sie entstanden unter Sauerstoffmangel in tieferen Gewässerzonen
durch den Abbau organischen Materials.
Auf diese Weise blieb die Verwesung aus und beim Abbau der organischen Materie entstanden Kohlenwasserstoffe.
Besonders bedeutend ist in dieser Hinsicht die Grube Messel nahe Darmstadt, Deutschland, deren Ölschiefer sich vor 47 Mio. Jahren und
über einen Zeitraum von 1,5 Mio. Jahren in einem rund 300 m tiefen Maar bildeten. Neben ausgezeichnet erhaltenen Vögeln, Säugetieren, Reptilien und Fischen sind auch fossile Käfer in den Messeler
Ölschiefern gefunden worden, die heute noch ihre prächtige Originalfärbung zeigen39 (s. Abb. 12). Prachtkäfer (Buprestidae) und Schnellkäfer (Elateridae) gehören zu den häufiger
in Ölschiefern nachgewiesenen Familien, deutlich seltener werden dagegen Bockkäfer (Cerambycidae) gefunden.
|
2.5 Neogen (23 mya bis heute) |
|
Während des Neogens nahmen die Kontinente schließlich ihre heutige Position ein. Der südamerikanische Subkontinent driftete nach Westen in Richtung der Subduktionszone
im Pazifik. Dabei wurden die Anden aufgefaltet. Während des Pliozän (5 mya) bildete sich eine Landbrücke zwischen Süd- und Nordamerika, über die der Faunenaustausch einsetzte.
Die Entstehung dieser Landbrücke hatte auch Auswirkungen auf das globale Klima. Der indische Subkontinent setzte seine Kollision mit Asien fort, bewegte sich jedoch zusätzlich westwärts, wodurch
der Kaukasus aufgefaltet wurde. Die Auffaltung des Himalaya dauert noch bis heute an.
Durch die Kollision von Afrika mit Europa und die Anhebung der Lithosphäre unter der Alborán See (westlichstes Mittelmeer) kam es zur Abschnürung des Mittelmeers
vom Atlantik. Während dieser Phase, die 600.000 Jahre von 6 bis 5,3 mya währte, trocknete das Mittelmeer fast ganz aus (Messinische Salinitätskrise). Erst am Ende der
Periode wurde das ausgetrocknete Becken durch einen engen, tiefen Kanal bei Gibraltar wieder geflutet, nach heutiger Ansicht zügig, aber nicht mit katastrophalen Auswirkungen.
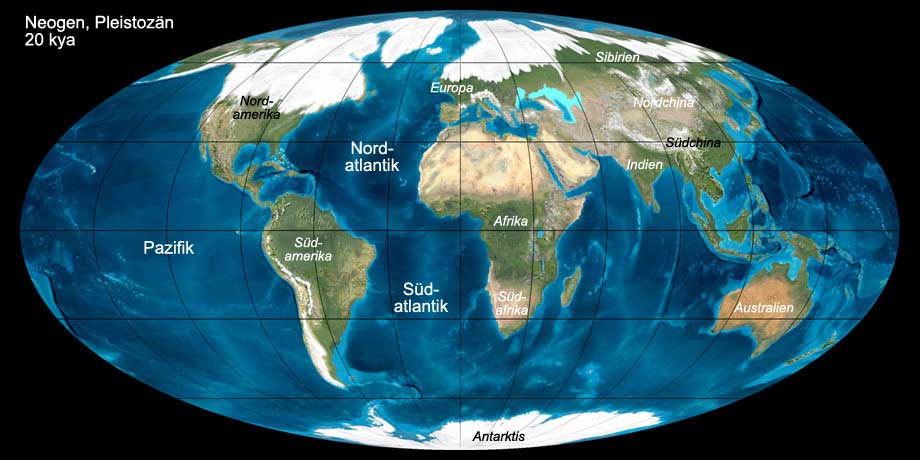
Abb. 13: Die Welt während des Neogens (Pleistozän, 20 kya). Anklicken, um Klima und Beschriftung ein-/auszublenden. © Ron Blakey, Verwendung mit schriftlicher Genehmigung.
| System |
Serie |
Stufe |
Alter (mya) |
| Neogen |
Holozän |
Holozän |
0,0117 - 0 |
| Pleistozän |
Jungpleistozän
(Tarantium) |
0,126 - 0,0117 |
Mittelpleistozän
(Ionium) |
0,781 - 0,126 |
Altpleistozän
(Calabrium) |
1,806 - 0,781 |
| Gelasium |
2,588 - 1,806 |
| Pliozän |
Piacenzium |
3,600 - 2,588 |
| Zancleum |
5,332 - 3,600 |
| Miozän |
Messinium |
7,246 - 5,332 |
| Tortonium |
11,608 - 7,246 |
| Serravallium |
13,82 - 11,608 |
| Langhium |
15,97 - 13,82 |
| Burdigalium |
20,43 - 15,97 |
| Aquitanium |
23,03 - 20,43 |
Das Neogen war eine Phase globaler Abkühlung, die schließlich in die Eiszeit des Pleistozäns mündete. Zu Beginn des Miozäns waren die Temperaturen auf der
nördlichen Halbkugel zunächst noch gemäßigt. Durch die Verbindung des süd- und des nordamerikanischen Kontinents im unteren Pliozän wurde die warme äquatoriale Meeresströmung
unterbrochen und es kam zu einer starken Abkühlung der Pole. Im Gelasium begannen sich die Eisschilde der Arktis und Antarktis zu bilden. Dies markierte den
Beginn eines neuen Eiszeitalters, das mit Wechseln von Kaltzeiten (Glaziale) und Warmzeiten (Interglaziale) bis heute anhält. Während der Glaziale dehnten sich die
Gletscher in einigen Gebieten bis zum 40. Breitengrad aus und bedeckten weite Teile Nordamerikas, Europas und Sibiriens. Durch die Eisbildung während der Kaltzeiten sank
der Meeresspiegel global um etwa 100 m ab, während der Interglaziale stieg er wieder an und überflutete die Küstenbereiche.
Die wichtigsten Fundstätten für fossile Käfer des Miozäns liegen in der warm-gemäßigten und subtropischen Zone. Viele Gattungen und Arten, die heute vorkommen,
existierten auch damals schon, allerdings unterschied sich ihre Verbreitung von der heutigen. Der wichtigste Fundort für Insekten des Pliozäns ist Willershausen
bei Göttingen mit hervorragend erhaltenen Käfern verschiedener Familien (Bock-, Rüssel-, Marienkäfer u.a.) sowie Vertretern anderer Insektenordnungen40.
In der Tongrube Willershausen wurden bisher 35 Gattungen aus 18 Käferfamilien nachgewiesen, davon sind heute sechs Gattungen ausgestorben.
Die pleistozäne Käferfauna ist sehr gut untersucht, beispielsweise in den Arbeiten von S. A. ELIAS41-43,
der die Zusammensetzung der Käferfauna zur Rekonstruktion der klimatischen Verhältnisse in den Rocky Mountains und auf Beringia, der ehemaligen Landbrücke
zwischen Asien und Nordamerika, heranzog.

Abb. 14: Pterolophosoma otiliae, Dominikanischer Bernstein, unteres Miozän44. © Entomapeiron / Francesco Vitali, Verwendung mit schriftlicher Genehmigung.
|
3. Zusammenfassung |
|
Aus entwicklungsgeschichtlicher Sicht ist die Ordnung der Käfer einzigartig. Heute sind 166 Familien bekannt, gegenüber 67 aus dem Paläozoikum und dem
Mesozoikum. 19 dieser Familien, darunter alle im Paläozoikum vorkommenden starben aus45 (vgl. Abb. 15). Viele rezente Familien lassen sich auf Vorfahren aus der Kreide, teilweise
auch aus dem Jura oder sogar der Trias, zurückführen - anders als bei vielen anderen Familien des Tierreichs, die sich nur bis zum Paläogen zurückverfolgen lassen. Mit anderen Worten bedeutet das,
dass die Entwicklung der Käfer gleichmäßiger und weniger rasant verlief als die anderer landbewohnender Tiergruppen.
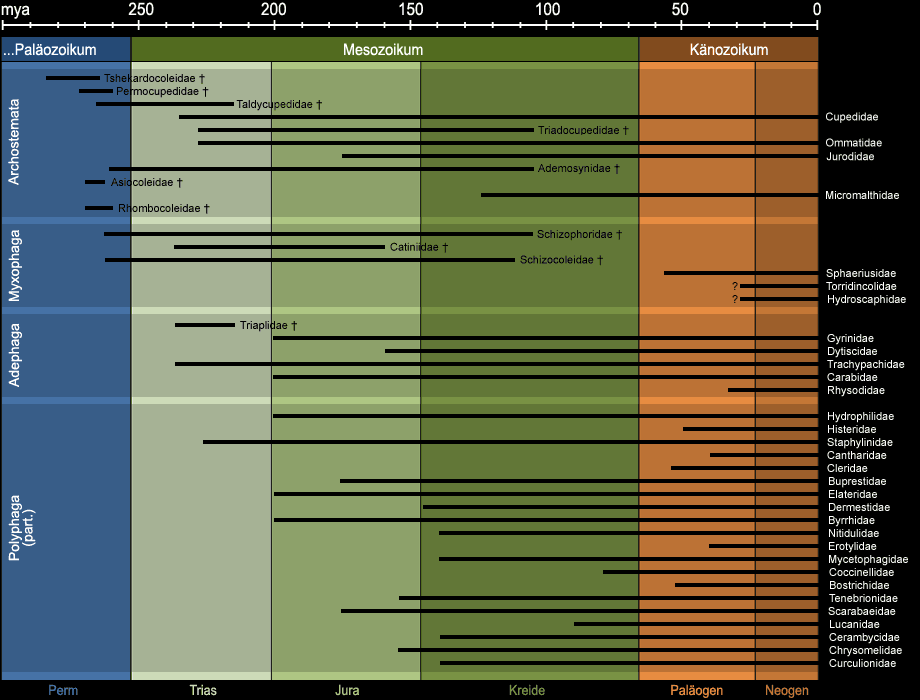
Abb. 15: Auftreten ausgewählter ausgestorbener und rezenter Käferfamilien in der Erdgeschichte. Die stark vereinfachte Darstellung wurde aus der
umfangreichen Literatur zum Thema zusammengetragen und erhebt keinen Anspruch auf Vollständigkeit.
A. P. VOGLER konstatierte 2007 in seiner bemerkenswerten Publikation über die Phylogenie der Käfer in Science2,
dass der evolutionäre Erfolg der Käfer weder auf die außerordentliche Diversifizierungsrate zurückzuführen ist, noch auf eine beherrschende Rolle als Pflanzenfresser
während des Aufstiegs der Bedecktsamer (Angiospermae) in der Kreide. Vielmehr legt der vorkreidezeitliche Ursprung von über 100 heutigen Abstammungslinien der
Käfer nahe, dass ihr Artenreichtum auf einer hohen Überlebensrate der Abstammungslinien und ihrer fortwährenden Diversifizierung in einem breiten Spektrum
ökologischer Nischen beruht.
Vielleicht gibt das (vermutlich apokryphe) Zitat des Genetikers und Evolutionsbiologen J. B. S. HALDANE (1892-1964) die gesuchte Antwort
auf die Frage, warum es so viele Käferarten gibt. HALDANE soll auf die Frage, welche Schlüsse man über Gott aus der Schöpfung ziehen könne, geantwortet haben:
"Eine übermäßige Vorliebe für Käfer".
|
Dank |
|
Die Erstellung dieser Themenseite wäre ohne Unterstützung mit Bildmaterial durch zahlreiche Wissenschaftler aus Paläontologie und Geologie nicht möglich gewesen.
Die zur Illustration gezeigten Fotografien fossiler Käfer sowie die Weltkarten wurden freundlicherweise von den nachgenannten Personen zur Verfügung gestellt,
denen ich hiermit herzlich danke:
- Ren Dong und Huali Chang, College of Life Science, Capital Normal University, Beijing, China
- Rafael G. Martins-Neto, Universidade Federal de Juiz de Fora, Juiz de Fora, Brasilien
- Oscar F. Gallego, Universidad Nacional del Nordeste, Corrientes, Argentinien
- Sonja Wedmann, Forschungsinstitut Senckenberg, Forschungsstation Grube Messel, Messel, Deutschland
- Carsten Brauckmann und Elke Gröning, Institut für Geologie und Paläontologie, TU Clausthal, Deutschland
- Roy Beckemeyer, Museum Associate of the University of Kansas Natural History Museum, Lawrence, KS, USA
- Ron Blakey, Northern Arizona University, Department of Geology, Flagstaff, AZ, USA
- Francesco Vitali, Genua, Italien
- Doug Lundberg, Amberica West, USA
Für die Durchsicht des Seitenentwurfs geht ein besonderer Dank an Irina Würtele und Klaas Reißmann.
|
Literaturverzeichnis
- Die Insektentaphozönose von Niedermoschel (Asselian, unt. Perm; Deutschland)
T. Hörnschemeyer, H. Stapf, Terra Nostra. Schriften der Alfred-Wegener-Stiftung 99/8: 98.
- On the systematic position of the supposed Permian beetles, Tshecardocoleidae [sic], with a description of a new collection from Moravia
J. Kukalová, Sb. Geol. Ved. Rada. P. Paleontol. 1969, 11, 139-161.
- A new family of Coleoptera from the Permian of the Urals
B. B. Rohdendorf, Dokl. Akad. Nauk. SSSR 1944, 44, 252-253.
- Paleozoic beetles of Cupedidae from the European USSR
A. G. Ponomarenko, Paleontol. Zh. 1963, 1, 70-85.
- Permocoleus, New Genus, the First Permian Beetle (Coleoptera) from North America
S. H. Lubkin, M. S. Engel Ann. Entomol. Soc. Am. 2005, 98(1), 73-76. 
- A Second Specimen of Permocoleus (Coleoptera) from the Lower Permian Wellington Formation of Noble County, Oklahoma
R. J. Beckemeyer, M. S. Engel, Journal of the Kansas Entomological Society 2008, 81(1), 4-7.
- The First Beetles (Permosynidae, Coleoptera) from the Upper Tatarian of European Russia
A. G. Ponomarenko, Paleontological Journal 2003, 37(2), 170-173.
- New beetles (Insecta: Coleoptera) from the Late Permian of South Africa
A. G. Ponomarenko, M. B. Mostovski, African Invertebrates 2005, 46, 253-260.
- On Permian and Triassic Insect Faunas in Relation to Biogeography and the Permian-Triassic Crisis
D. E. Shcherbakov, Paleontological Journal 2008, 42(1), 15-31.
- Beetles (Insecta, Coleoptera) of the Late Permian and Early Triassic
A. G. Ponomarenko, Paleontological Journal 2004, 38 (Suppl. 2), S185-S196.
- New Triassic Beetles (Coleoptera) from Northern European Russia
A. G. Ponomarenko, Paleontological Journal 2008, 42(6), 600-606.
- Fossil insects from the Tithonian "Solnhofener Plattenkalke" in the Museum of Natural History, Vienna
A. G. Ponomarenko, Ann. Naturhist. Mus. Wien 1985, 87, 135-144.
- A New Genus of Elateriform Beetles (Coleoptera, Polyphaga) from the Middle-Late Jurassic of Karatau
E. V. Yan, Paleontological Journal 2009, 43 (1), 78-82.
- New Ommatids from the Late Jurassic of western Liaoning, China (Coleoptera: Archostemata)
J.-J. Tan, D. Ren, M. Liu, Insect Science 2005, 12, 207-216.
- New Cupedids from the Middle Jurassic of Inner Mongolia, China (Coleoptera: Archostemata)
J.-J. Tan, D. Ren, C. Shih, Annales Zoologici (Warszawa) 2006, 56(1), 1-6.
- Palaeogeography, Palaeoecology and Taphonomy of Jurassic-Cretaceous Cupedomorpha Faunas from China
J.-J. Tan, D. Ren, C. Shih, Mesozoic Terrestrial Ecosystems 2006, 130-136.
- New Beetles of the Family Cupedidae from the Mesozoic of Mongolia. Ommatini, Mesocupedini, Priacmini
A. G. Ponomarenko, Paleontological Journal 1997, 31(4), 389-399.
- Early Jurassic Insects from the Newark Supergroup, Northeastern United States
P. Huber, N. G. McDonald, P. E. Olsen in P. M. LeTourneau and P.E. Olsen (eds.), The Great Rift Valleys of Pangea in Eastern
North America, 2003, Volume 2: Sedimentology, Stratigraphy, and Paleontology, Columbia University Press, New York, 206-223.
- Insect diversity in the fossil record
C. C. Labandeira, J. J. Sepkoski, Science 1993, 261, 310-315.
- The fossil record of weevils and related beetle families (Coleoptera, Curculionoidea)
V. G. Gratshev, V. V. Zherikhin, Acta zoologica cracoviensia 2003, 46, 129-138.
- A new genus and two new species of fossil Elaterids from the Yixian Formation of Western Liaoning, China (Coleoptera: Elateridae)
H. Chang, F. Zhang, D. Ren, Zootaxa 2008, 1785, 54-62.
- New Fossil Elaterids (Coleoptera: Polyphaga: Elateridae) From the Jehol Biota in China
H. Chang, A. Kirejtshuk, D. Ren, Annals of the Entomological Society of America, 2010, 103(6), 866-874.
- A Survey of Mesozoic Buprestids (Coleoptera) from Eurasian deposits
A. V. Alexeev, Proceedings of the First International Palaeoentomological Conference, Moscow 1998.
- On Mesozoic Buprestids (Coleoptera: Buprestidae) from Russia, Kazakhstan, and Mongolia
A. V. Alexeev, Paleontological Journal 2000, 34(3), S323-S326.
- Jurassic and Lower Cretaceous Buprestidae (Coleoptera) from Eurasia
A. V. A. Orekhovo-Zuyevo, Paleontological Journal 1993, 27(1A), 9-34.
- The Crato Fossil Beds of Brazil - Window into an Ancient World
D. M. Martill, G. Bechly and R. F. Loveridge, Cambridge University Press 2007, ISBN 978-0-521-85867-0.
- New beetles (Insecta: Coleoptera) from the Lower Cretaceous of Spain
A. Ponomarenko, X. Martínez-Delclòs, Acta Geologica Hispanica 2000, 35(1-2), 47-52.
- New Jewel Beetles (Coleoptera: Buprestidae) from the Cretaceous of Russia, Kazakhstan, and Mongolia
Paleontological Journal 2009, 43(3), 277-281.
- The Role of Insects in Late Jurassic to Middle Cretaceous Ecosystems
C. C. Labandeira in S. G. Lucas, J. I. Kirkland and J. W. Estep (eds.), 1998, Lower and Middle Cretaceous
Terrestrial Ecosystems, New Mexico Museum of Natural History and Science Bulletin No. 14., 105-124.
- Dinosaurs, dung beetles, and conifers; participants in a Cretaceous food web
K. Chin, and B. D. Gill, Palaios 1996, 11(3), 280-285.
- Did dinosaurs have any relation with dung-beetles? (The origin of coprophagy)
A. Arillo, V. M. Ortuño, Journal of Natural History 2008, 42(19&20), 1405-1408.
- Timing the Radiations of Leaf Beetles: Hispines on Gingers from Latest Cretaceous to Recent
P. Wilf, C. C. Labandeira, W. J. Kress, C. L. Staines, D. M. Windsor, A. L. Allen, K. R. Johnson, Science 2000, 289, 291-294.
- A New Weevil-Beetle (Insecta, Coleoptera, Nemonychidae) from the Lower Cretaceous of Spain
V. V. Zherikhin and V. G. Gratshev, Paleontological Journal 2003, 37(4), 407-408.
- Fossil Curculionoid Beetles (Coleoptera, Curculionoidea) from the Lower Cretaceous of Northeastern Brazil
V. V. Zherikhin and V. G. Gratshev, Paleontological Journal 2004, 38(5), 528-537.
- New fossil dermestid beetles (Coleoptera: Dermestidae) from the Baltic amber - II
J. Háva, J. Prokop, M. Kadej, Studies and reports of District Museum Prague-East Taxonomical Series 2, 2006, 1-2, 65-68.
- New fossil dermestid beetles (Coleoptera: Dermestidae) from the Baltic amber - III
J. Háva, J. Prokop, A. Herrmann, Acta Soc. Zool. Bohem. 2007, 71, 151-157.
- Osoriinae (Coleoptera: Staphylinidae) from Dominican Amber
U. Irmler, Stuttgarter Beitr. Naturk. Ser. B 2003, 342, 1-16.
- New Amber Deposit provides evidence of early Paleogene extinctions, paleoclimates, and past distibutions
G. Poinar Jr., B. Archibald, A. Brown, The Canadian Entomologist 1999, 131, 171-177.
- Fossile Prachtkäfer (Coleoptera: Buprestidae: Buprestinae) aus dem Mitteleozän der Grube Messel bei Darmstadt, Teil 1
T. Hörnschemeyer, S. Wedmann, Cour. Forsch.-Inst. Senckenberg 1994, 170, 85-136.
- Dritter Beitrag über Käfer (Coleoptera) aus dem Jungtertiär von Willershausen, Bl. Northeim 4226
E. Gersdorf, Geol. Jahrb. A 1976, 36, 103-145.
- Late Pleistocene and Holocene Seasonal Temperatures Reconstructed from Fossil Beetle Assemblages in the Rocky Mountains
S. A. Elias, Quaternary Research 1996, 46 (3), 311-318.
- Late Pleistocene Climates of Beringia, Based on Analysis of Fossil Beetles
S. A. Elias, Quaternary Research 2000, 53 (2), 229-235.
- Climatic Tolerances and Zoogeography of the Late Pleistocene Beetle Fauna of Beringia
S. A. Elias, Géographie physique et Quaternaire 2000, 54 (2), 143-155.
- A new cerambycid from Dominican amber and remarks on the fossil Plectromerus-species (Coleoptera Cerambycidae)
F. Vitali, Entomapeiron Paleoentomology 1(1): 1-12
- The geological history of beetles
A. G. Ponomarenko in J. Pakaluk, S. A. Ślipiński (eds.), 1995, Biology, phylogeny, and classification of Coleoptera:
papers celebrating the 80th birthday of Roy A. Crowson, 155-171. Museum i Instytut Zoologii PAN, Warszawa.
|
Weiterführende Literatur
- Evolution and Classification of Beetles
J. F. Lawrence, A. F. Newton Jr., Ann. Rev. Ecol. Syst. 1982, 13, 261-290.
- A Comprehensive Phylogeny of Beetles Reveals the Evolutionary Origins of a Superradiation
A. P. Vogler et al., Science 2007, 318, 1913-1916.
- "Inordinate Fondness" Explained: Why Are There So Many Beetles?
B. D. Farrell, Science 1998, 281, 555-559.
- Fossil insects in Gondwana - localities and palaeodiversity trends
T. Schlüter, Acta zoologica cracoviensia 2003, 46, 345-371.
- Ecological evolution of beetles (Insecta: Coleoptera)
A. G. Ponomarenko, Acta zoologica cracoviensia 2003, 46, 319-328.
- Evolutionary Rates in the Adaptive Radiation of Beetles on Plants
B. D. Farrell, A. S. Sequeira, Evolution 2004, 58(9), 1984-2001.
- The Evolution of Agriculture in Beetles (Curculionidae: Scolytinae and Platypodinae)
B. D. Farrell et al., Evolution 2001, 55(10), 2011-2027.
|

 Abb. 12: Der Prachtkäfer Psiloptera weigelti, Grube Messel, Deutschland, 47 mya. © Forschungsinstitut Senckenberg, Verwendung mit schriftlicher Genehmigung.
Abb. 12: Der Prachtkäfer Psiloptera weigelti, Grube Messel, Deutschland, 47 mya. © Forschungsinstitut Senckenberg, Verwendung mit schriftlicher Genehmigung.











